Estructura y diversidad de la
vegetación leñosa en un área de matorral espinoso tamaulipeco con actividad
pecuaria en Nuevo León, México
Structure and diversity of the woody vegetation in a Tamaulipas thornscrub with livestock activity in Nuevo León,
Mexico
Tania Isela Sarmiento-Muñoz1,2 , Eduardo Alanís-Rodríguez1 , José
Manuel Mata-Balderas2 *, Arturo Mora-Olivo3
*Correspondencia:
manuelmata792@gmail.com/Fecha de recepción: 13 de octubre de 2017/Fecha de
aceptación: 5 de abril de 2019/Fecha de publicación: 29 de julio de 2019.
1Universidad
Autónoma de Nuevo León, Facultad de Ciencias Forestales, carretera Linares-Cd.
Victoria km 145, Apartado Postal 41, Linares, Nuevo León, México, C. P. 67700.2
Gestión Estratégica y Manejo Ambiental, S. C. 3Universidad Autónoma
de Tamaulipas, Instituto de Ecología Aplicada.
RESUMEN
En el noreste de México
existe un incremento en el establecimiento de pastizales para la ganadería, lo
cual ha ocasionado cambios en la estructura y diversidad florística. El
objetivo de la presente investigación fue determinar algunos parámetros dasométricos y de diversidad de los elementos leñosos
presentes en un área de matorral espinoso tamaulipeco (MET), con actividad
pecuaria, en el estado de Nuevo León, México, para establecer si las
modificaciones realizadas contribuyen o no a la conservación de la diversidad
vegetal. La actividad pecuaria que se realizaba en el área de estudio era un
aprovechamiento mixto de moderada intensidad, con aclareos por lo bajo (d ˂ 5
cm). Se seleccionaron cuatro sitios de muestreo de 1 600 m2 cada
uno. Se recolectaron datos sobre variables dasométricas
y se obtuvieron índices ecológicos. Los resultados permitieron un registro de 8
familias, 16 géneros y 17 especies arbóreas y arbustivas. La densidad absoluta
reportada fue de 309 individuos/ha, mientras que el área de copa estimada fue
de 9 521.83 m2 /ha. La riqueza (Da = 3.03) y diversidad (H´ = 2.31)
específicas, resultaron en valores similares a matorrales sin actividad
productiva. Las especies con mayor importancia dentro del área de estudio
fueron características del MET: Prosopis laevigata (IVI = 28.59), Havardia pallens (IVI = 13.41) y Cordia boissieri
(IVI = 8.89), de las cuales, Prosopis laevigata tiene antecedentes de uso forrajero. Se
concluye que la composición de especies arbóreas y arbustivas en este sistema
pecuario conserva atributos deseables de las comunidades vegetales nativas,
como la riqueza de especies, diversidad, densidad y área de copa.
PALABRAS CLAVE: aclareo,
riqueza, ganadería, atributos de conservación.
ABSTRACT
In
the northeastern region of Mexico, there is an increase in the establishment of
grasslands for livestock, which has created changes in the structure and
floristic diversity of ecosystem. The aim of this study was to determine the
structure and diversity of the woody species ve- getation of an area of Tamaulipas thornscrub
(MET) which was modified for livestock rising in Nuevo
Leon, Mexico, to establish whether or not the changes have contributed to the
conservation of plant diversity. The livestock activity practiced in the
researched area is mixed purpose use at a moderate intensity, with a thinning
treatment at the lower vegetation layer (d ˂ 5 cm) to create open areas for
cattle grazing in the study area. Four sampling plots of 1 600 m2
each were selected. Dasometric measures were recorded
and ecological indexes were estimated. The results are a record of 8 families,
16 genus and 17 species of trees and shrubs. The absolute density was 309/ha,
while the estimated canopy area was 9 521.83 m2 /ha. Richness (Da =
3.03) and species diversity (H’ = 2.31) resulted in similar values to scrubs
with nonproductive activity. Species with the greatest IVI are characteristic
of the MET: Prosopis laevigata
(IVI = 28.59), Havardia pallens (IVI
= 13.41) and Cordia boissieri
(IVI = 8.89), from which, Prosopis laevigata is considered as a species for forage use. We
conclude that the composition of the tree and shrub species in this livestock
rising system contributes to preserving the desirable attributes of the native
vegetation communities, such as richness and species diversity, density and
canopy area.
KEYWORDS:
thinning, richness, livestock use, conservation attributes.
INTRODUCCIÓN
Estudios recientes en México
indican que, el factor de mayor influencia en la degradación de los ecosistemas
y la biodiversidad, es el cambio del uso de la tierra (Sarukhán
y col., 2015). De igual forma, la destrucción del hábitat y la fragmentación
han tenido un gran impacto en los ecosistemas terrestres y acuáticos
(Williams-Linera, 2002; Santos y Tallería, 2006;
Rodríguez-Estrella y col., 2016). Por ejemplo, en 1976, la cobertura original
de la vegetación primaria terrestre en el país se redujo en un 54 %, mientras
que para 2011, la extensión original de la vegetación cubierta, en su condición
primaria y secundaria, se menguó a 72 % según información del Instituto
Nacional de Estadística y Geografía (INEGI, 2013). Sin embargo, recientemente,
las tasas de deforestación han disminuido en muchas regiones del país, en parte
debido a que las áreas restantes son inaccesibles, consideradas de bajo valor
comercial o bien se encuentran bajo algún régimen de protección (Rosete-Vergés y col., 2014; FAO, 2016). Más allá de esto,
actualmente, una parte significativa de la vegetación restante se compone de
pequeños fragmentos aislados o está representada por el crecimiento secundario,
es decir, vegetación en proceso de regeneración, después de ser sometida a
perturbación de acuerdo a la Comisión Nacional para el Conocimiento y Uso de la
Biodiversidad (CONABIO, 2009).
Un factor importante en el
cambio del uso del suelo es la actividad pecuaria (ArriagaCabrera,
2009). De las 1 960 189 ha del territorio continental, a nivel nacional,
aproximadamente 81 758 631 ha se identifican con uso de agostadero, enmontado o
de otro tipo (INEGI, 2017). Esta situación se refleja en el norte de México,
con el constante incremento de las actividades pecuarias y agrícolas (Návar-Chaidez, 2008; Cantú-Silva y col., 2017). Dichas
prácticas han ocasionado cambios en la estructura del ecosistema y en su
diversidad florística, con la consecuente pérdida de especies y material
genético, invasión de especies generalistas y la alteración de las funciones,
como la polinización y dispersión de semillas, entre otros (Jiménez-Pérez y
col., 2012; Moreno-Medina y col., 2017).
Actualmente se busca integrar
el uso y la conservación de los recursos naturales con las necesidades
productivas de la población (Maass y col., 2005; Jose, 2012; SAGARPA, 2013). Desde hace 30 años, en el sector
agropecuario, han surgido prácticas, en las que se integra y combina el manejo
de árboles o arbustos con ganado (Musálem, 2002;
Sánchez y col., 2003). Constituyen actividades agroforestales, que se han
empezado a utilizar por ganaderos, con el fin de incrementar la productividad
de sus tierras y promover la conservación de los recursos naturales (CONAFOR,
2010; SAGARPA, 2013).
Los sistemas agroforestales
brindan beneficios múltiples, como la dispersión de especies vegetales y la
conservación de animales silvestres y el paisaje conjunto (Mahecha y Zoot, 2002; Barrance y col.,
2009). En términos de productividad, se ha descrito que los árboles presentes
en ellos, además de proveer madera para cercas y combustible, generan un
microclima mediante la sombra, que el ganado aprovecha (Mitlohner
y col., 2002). Estas prácticas han sido tema de estudio, principalmente en los
trópicos y ecosistemas templados del continente y del país, enfocándose
primordialmente a la productividad de los mismos (Sánchez y col., 2003; Ramírez
y col., 2012; Rosales-Adame y col., 2014). Aunque, se ha preconcebido la
importancia y el potencial ecológico que sostienen las especies arbóreas que
los integran, la evidencia científica que comprueba estas afirmaciones es aún
escasa, en particular en las comunidades vegetales áridas y semiáridas.
El matorral espinoso
tamaulipeco (MET) es una comunidad vegetal semiárida, caracterizada por la
presencia de una alta diversidad de plantas, con elevados valores de densidad
en su estrato arbóreo-arbustivo (Cram y col., 2006;
Alanís-Rodríguez y col., 2008; González-Rodríguez y col., 2010;
Alanís-Rodríguez y col., 2015; Pequeño-Ledezma y col., 2017). Sin
embargo, es una de las
comunidades en donde la ganadería extensiva ha sido practicada durante los últimos
350 años (Foroughbakhch y col., 2009), siendo esta
una de las causas principales de su deforestación (Arriaga-Cabrera, 2009;
Alanís-Rodríguez y col., 2015), por lo que resulta importante encontrar y
aplicar nuevas prácticas de aprovechamiento que conserven las características
mencionadas. En el MET se han reportado aproximadamente 80 especies maderables,
de las cuales, 9 han sido destacadas con potencial agroforestal (Pando-Moreno y
Villalón-Mendoza, 2001).
El objetivo de esta
investigación fue evaluar algunos parámetros dasométricos
y de diversidad de los elementos leñosos presentes en un matorral espinoso
tamaulipeco (MET), sometido a la actividad pecuaria en el estado de Nuevo León,
ubicado en la región noreste de México, para determinar si las modificaciones
realizadas para este uso, contribuyen o no, a la conservación de la diversidad
vegetal, con base en estudios recientes del MET.
MATERIALES
Y MÉTODOS
Área
de estudio
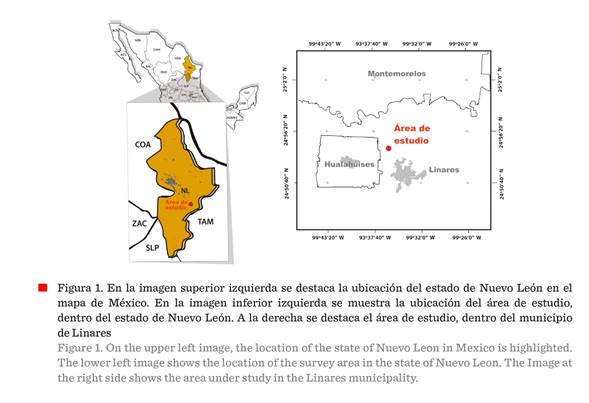
El área se ubica en el ejido
Emiliano Zapata, municipio de Linares, Nuevo León, en el noreste de México
(Figura 1). Las coordenadas registradas están dentro del rango de los 24°54’ N
y los 99°35’ W, en el suroeste de la región fisiográfica Llanura Costera del
Golfo Norte (INEGI, 1986). El clima dominante en esta región es semicálido subhúmedo, con precipitación mayor a los 800 mm. El sistema de topoformas
pertenece a lomerío suave con bajada, con predominancia de suelos regosoles. Las posibilidades de uso pecuario son de
pastoreo extensivo sobre vegetación natural distinta del pastizal (INEGI,
1986).
La vegetación principal es el
MET, en el cual predominan las asociaciones de estratos arbustivos altos o
medianos y arbóreos altos, caracterizados en su mayoría por especies espinosas,
con hojas compuestas (García-Hernández y Jurado, 2008; Alanís-Rodríguez y col.,
2013). Las especies más representativas son Castela
texana (Torr. & A. Gray) y Rose, Celtis pallida Torr., Cordia boissieri A. DC., Ebenopsis ebano (Berland.)
Barneby
& J.W. Grimes,
Parkinsonia texana var. macra (I.M. Johnst.) Isely, Prosopis laevigata (Humb. & Bonpl. ex Wild.) M.C. Johnst., Randia obcordata S.
Watson, Sideroxylon celastrinum (Kunth) T.D. Penn. y Vachellia rigidula (Benth.) Seigler & Ebinger, entre otras (Alanís-Flores, 1996; Jiménez-Pérez y
col., 2013).
Análisis
de la vegetación
En una superficie de 19 249 m2
se realizaron aclareos por lo bajo en los años 2011 y 2012, eliminando la
mayoría de los individuos con d < 5 cm. La intención de este manejo fue
modificar la estructura de la comunidad vegetal existente, creando espacios
abiertos para el establecimiento de pastos y permitir el desplazamiento del
ganado. Sin embargo, se conservan los árboles y arbustos de diámetros mayores.
Las copas de los árboles brindan sombra que favorece el resguardo del ganado en
temporadas de calor y proporcionan una fuente de alimento para el ganado y las
aves, en variadas épocas del año.
En junio del año 2013, se
realizó la evaluación de la comunidad vegetal, mediante el establecimiento de 4
sitios de muestreo de 1 600 m2 cada uno (33 % del área de estudio),
distribuidos al azar. En los sitios de muestreo se evaluaron todas las especies
arbóreas y arbustivas. Se registró la nomenclatura taxonómica de cada individuo
y se realizaron mediciones de altura total (h), diámetro basal (d0.5) y
cobertura de copa, en sentido nortesur y este-oeste.
El d0.5 se midió a
partir de las bifurcaciones que presentaron los individuos a mayor altura, ya
que resulta más práctico y económico realizar una sola medición
(Alanís-Rodríguez y col., 2008).
Análisis
de la información
Para evaluar el significado
ecológico de las especies arbóreas se estimaron los indicadores relativos de
densidad (ARi),
dominancia (DRi),
frecuencia (FRi)
e índice de valor de importancia (IVI)
(Müeller-Dombois y Ellenberg,
1974; Magurran, 2004), calculados a partir de las
siguientes ecuaciones. La densidad relativa se obtuvo por medio de la densidad
absoluta, y el cálculo de su valor relativo. Se utilizaron las siguientes
ecuaciones:
![]()
Donde: Ai es la densidad absoluta de la especie
i, Ni es el número de individuos de la especie i, y S la superficie muestral en ha.
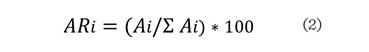
Donde: ARi es la
densidad relativa de la especie i, Ai con respecto a la densidad total ∑Ai.
Para estimar la dominancia
relativa también se empleó el uso de dos ecuaciones, la dominancia absoluta y
el cálculo de su valor relativo. Se utilizaron las siguientes ecuaciones:
![]()
Donde: Di es la dominancia absoluta de la especie i, Abi
el área basal de especie i, y S la superficie muestral
(ha).
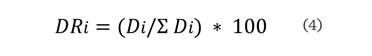
Donde: DRi es la
dominancia relativa de la especie i, Di con respecto a la dominancia total ∑Di
La frecuencia relativa se
obtuvo calculando la frecuencia absoluta y posteriormente su valor relativo. Se
utilizaron las siguientes ecuaciones:
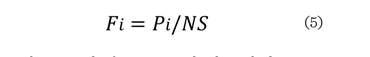
Donde: Fi es la frecuencia absoluta de la especie i, Pi es la frecuencia de
la especie i en los sitios de
muestreo, y NS el número total de
sitios de muestreo.
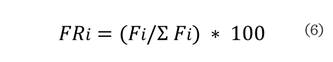
Donde: FRi es la
frecuencia relativa de la especie i, con respecto a la frecuencia total ∑Fi. El índice de valor de importancia (IVI) se define como:
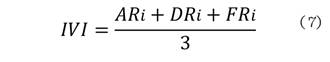
Donde: ARi es la
densidad relativa de la especie i, DRi es la
dominancia relativa de la especie i,
y FRi es la
frecuencia relativa de la especie i.
Para estimar la diversidad
alfa y la riqueza de especies se utilizaron, el índice de ShannonWiener
(H´) (Shannon y Weaver, 1949) y el índice de Margalef (Clifford y Stephenson, 1975), respectivamente. El índice de
Shannon-Wiener se estimó mediante la siguiente ecuación:
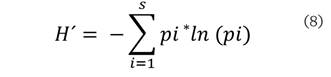
Donde: S es el número de especies presentes, ln es logaritmo natural y pi
es la proporción de las especies, la cual se calcula con la siguiente ecuación:
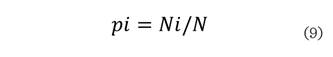
Donde: Ni es el número de individuos de la especie i y N es el número total
de individuos.
Con el mismo significado de
las variables en común, el índice de diversidad de Margalef
(DMg) se estimó con la ecuación:
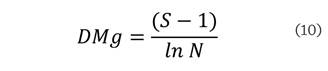
La distribución de las
especies en tres diferentes zonas de altura se calculó mediante el índice de
distribución vertical de Pretzsch (Pretzsch, 1996), el cual toma tres zonas de altura: zona I:
80 a 100 % de la altura máxima de la población; zona II: 50 % a 80 %; zona III:
0 % a 50 %.
Posteriormente, se calculó el
índice de distribución vertical de especies (A), el cual cuantifica la diversidad de especies y su ocupación en
el espacio vertical en la población (Estrada-Castillón y col., 2005). El índice
de distribución vertical se calculó de acuerdo a la siguiente fórmula:
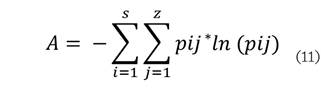
Donde: S es el número de especies presentes; Z el número de zonas de altura y pij la proporción de especies en
cada zona de altura, la cual se calcula con la siguiente ecuación:
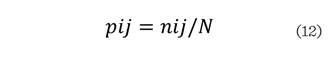
Donde: nij es el número
de individuos de la misma especie (i)
en la zona (j) y N el número total de
individuos.
Del índice A se derivan el Amax, que corresponde al valor
máximo de A, dado por el número de
especies (S) y zonas de altura (Z); y el Arel, que es la estandarización en porcentaje del índice A. Estos son indicativos de la
distribución de las especies en los estratos de altura, de manera que, entre
más cercano sea el valor de Amax al índice de A,
más se acerca el valor de Arel al 100
%, lo cual se presenta cuando el número total de individuos se encuentra
equitativamente distribuido entre estratos y especies (Rubio-Camacho y col.,
2014).
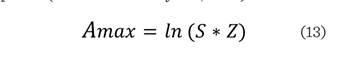
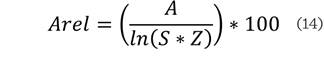
Para describir la estructura
de la comunidad en términos de la densidad de cada especie, se utilizó un
modelo que describe la relación gráfica entre el valor de la densidad (en una
escala logarítmica) y las especies, en función de un arreglo secuencial de la
más a la menos abundante (Martella y col., 2012).
RESULTADOS
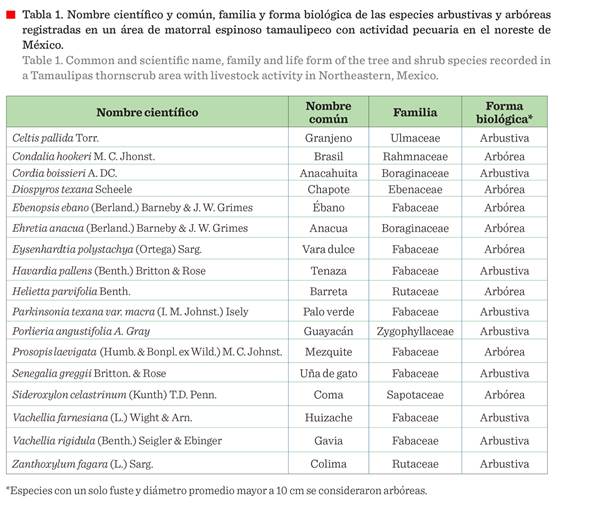
Se registraron 17 especies
arbóreas y arbustivas, distribuidas en 16 géneros y 8 familias de plantas
vasculares (Tabla 1). La familia con mayor riqueza específica fue Fabaceae, con 8 especies.
Parámetros
ecológicos
Densidad
La densidad absoluta, para el
total del área de estudio, fue de 309 individuos/ha. Las 3 especies más
abundantes fueron Prosopis leavigata (Humb. & Bonpl. ex Wild.) M. C. Johnst., Havardia pallens (Benth.) Britton & Rose y Celtis pallida Torr.
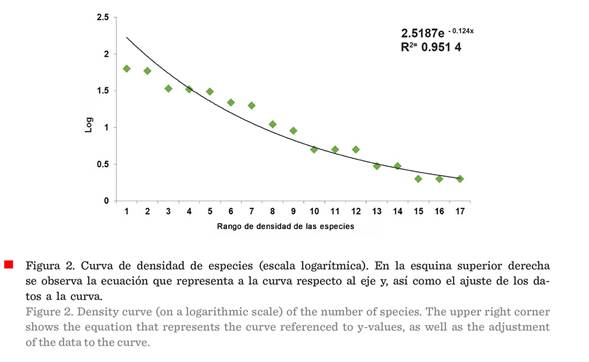
(Tabla 2). La forma de la curva de la densidad de las especies (en una escala
logarítmica) fue una Log-normal típica, con una línea de tendencia exponencial,
la cual es muy útil cuando los valores de los datos aumentan o disminuyen a
intervalos cada vez mayores (Figura 2). Se observa que el valor R2
es 0.951 4, cercano a 1, lo cual indica un buen ajuste de la curva a los datos.
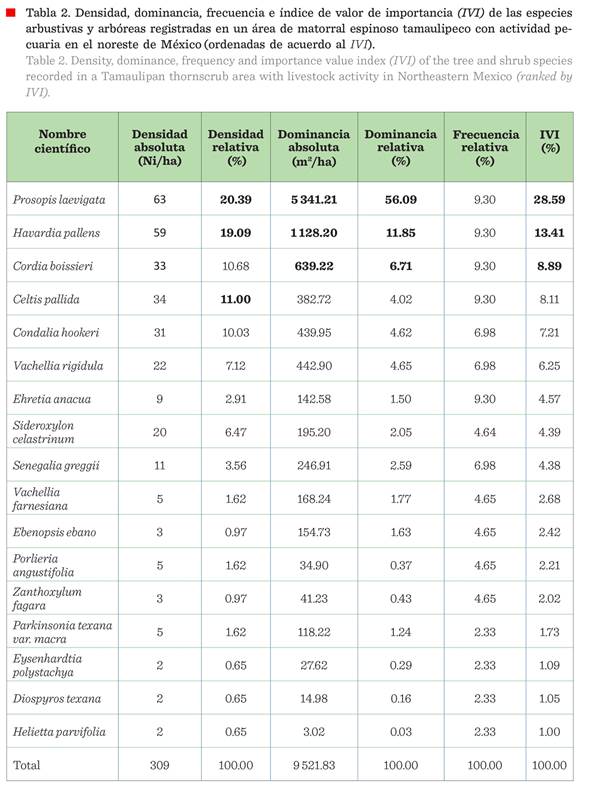
Dominancia
La cobertura de copa en el
área fue de 9 521.83 m2 /ha, lo que indica que, en general, existe
un 95 % de la superficie cubierta por el dosel, y únicamente el 5 % se
encuentra desprovista (asumiendo mínimo traslape de copas). Prosopis leavigata fue la especie dominante, con
56.09 % de la cobertura de copa de todas las especies en el área de estudio. La
segunda especie en dominancia fue Havardia pallens, seguida de Cordia boissieri.
Frecuencia
Se registró la presencia de 5
especies en todos los sitios de muestreo, Celtis pallida, Cordia boissieri, Ehretia anacua (Berland.) Barneby & J. W. Grimes, Harvardia pallens y Prosopis leavigata,
cada una con 9.3 % de frecuencia relativa (Tabla 2). Le siguen Condalia hookeri M. C.
Jhonst., Vachellia rigidula y Senegalia greggii Britton. & Rose,
con presencia en cuatro de los 5 sitios de muestreo.
Índice
de valor de importancia (IVI)
La especie con mayor peso
ecológico, según el índice de valor de importancia fue Prosopis leavigata (28.59 %), le siguen Havardia pallens y Cordia boissieri.
Las especies con menor peso ecológico fueron Helietta parvifolia Benth.
y Diospyros texana Scheele (Tabla 2).
Distribución
vertical de las especies
Las especies del estrato I
registraron una densidad de 16 individuos/ha, del estrato II de 208
individuos/ha y del estrato III de 85 individuos/ha; lo que indicó que la
comunidad vegetal era cohortal, es decir, que
predominaba un solo estrato (II) (Figura 3). De acuerdo al índice de distribución
vertical de especies (A), el estrato I (alto, de 9.85 m a 12.3 m) se encontraba
conformado por las especies Prosopis leavigata, Havardia pallens y Vachellia farnesiana (L.) Wight & Arn.
En el estrato II (medio, de 6.16 m a 9.84 m), que fue el mejor representado,
más del 50 % de las especies fueron Havardia pallens, Prosopis laevigata, Condalia hookeri y Vachellia rigidula, entre otras, con menor presencia. Havardia pallens fue la especie
predominante en este estrato. Por otro lado, el estrato III (bajo, menor a 6.16
m) se encontraba conformado casi en un 60 % por Cordia boissieri, Celtis pallida y Sideroxylon celastrinum, entre otras, con menor
presencia; Sideroxylon celastrinum alcanzó
la altura máxima de 6.14 m.
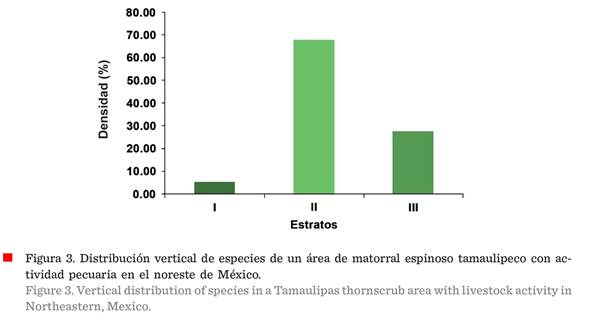
El índice de distribución
vertical (A) fue de 2.88, con un Amax de 3.93 y un Arel de 73 %, lo que indica uniformidad alta, en cuanto a la
diversidad de altura.
Densidad
de los individuos por clase diamétrica
Al analizar la densidad de
individuos por ha, de acuerdo a las clases diamétricas,
mediante un gráfico de barras, se observó una línea logarítmica, con tendencia
negativa en la densidad de individuos, conforme aumenta el diámetro de los
mismos (Figura 4). Las densidades mayores de individuos ( ≥
60 individuos/ha), se concentraron en las clases diamétricas
I y II.
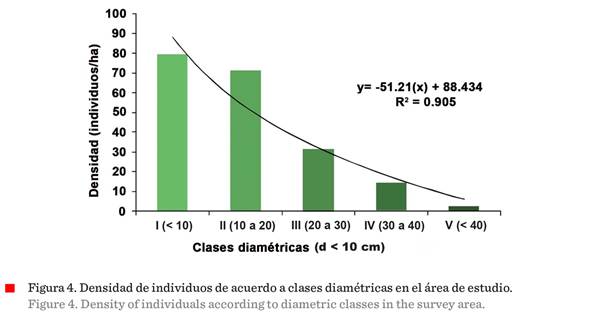
Diversidad
El índice de Margalef (DMg) fue de 3.03 y el índice de Shannon–Wiener (H´) de 2.31.
DISCUSIÓN
La familia con mayor número
de especies fue Fabaceae, lo cual coincide con
estudios recientes realizados en el MET, bajo diversas condiciones
(Estrada-Castillón y col., 2005; González-Rodríguez y col., 2010; Jiménez-Pérez
y col., 2012; Molina y col., 2013; Mora-Donjuán y col., 2013a; 2013b;
Pequeño-Ledezma y col., 2017). Comparando con otras evaluaciones del MET, en donde
se aplicó la misma metodología (igual número y dimensiones de los sitios de
muestreo), la riqueza específica registrada fue mayor, que en áreas con
regeneración pasiva e historial de pastoreo intensivo (Pequeño-Ledezma y col.,
2012; Yerena-Yamallel y col., 2014), pero menor que
en áreas sin disturbio reciente (Alanís-Rodríguez y col., 2011; Mora-Donjuán y
col., 2013a; Yerena-Yamallel y col., 2014).
Parámetros
ecológicos
Densidad Havardia pallens,
la segunda especie de mayor densidad relativa, en este estudio, ha sido
reportada como una de las tres especies con mayor densidad relativa en áreas de
MET sin disturbio (García-Hernández y Jurado, 2008; Yerena-Yamallel
y col., 2014) y en terrenos con historial de ganadería extensiva
(Pequeño-Ledezma y col., 2012). Por el contrario, Prosopis leavigata y Celtis pallida, las cuales fueron también dos
de las especies más abundantes en este trabajo, presentaron menor densidad en
los estudios referidos, así como en el trabajo de Jiménez-Pérez y col. (2013),
quienes analizaron áreas del MET con diferentes tipos de alteraciones.
La forma de la curva de
densidad estuvo determinada por la densidad por especie, la cual sigue una
distribución normal (Figura 2), con muchas especies con valores intermedios y
pocas con valores mínimos o máximos de densidad. Este tipo de distribución se
encuentra en la mayoría de las comunidades vegetales (Martella
y col., 2012), como el MET en el presente estudio, donde las especies Prosopis leavigata y Havardia pallens
fueron las que dominaron con valores máximos de densidad, mientras que Eysenhardtia polystachya
(Ortega) Sarg., Diospyros texana y Helietta parvifolia
registraron los valores mínimos (Tabla 2).
Dominancia
Frecuentemente, las especies
reportadas como más dominantes en el MET, con historial pecuario y agrícola,
son Vachellia farnesiana y Vachellia rigidula
(Alanís-Rodríguez y col., 2008; Pequeño-Ledezma y col., 2012; JiménezPérez y col., 2013; Mora-Donjuán y col., 2013b);
Diferente a lo que reportaron Mora-Donjuán y col. (2013a; 2013b) en un área sin
disturbio reciente, donde las especies dominantes fueron Vachellia amentacea, Diospyros texana y Cordia boissieri,
con esta última presente en el sistema pastoreado. La suma total de la
dominancia absoluta de las especies en el área de estudio (Tabla 2) es mayor a
los registros de dominancia absoluta documentadas por Mora-Donjuán y col.
(2013b), en sus tres áreas de estudio del MET (dos con historial pecuario y una
sin disturbio).
Frecuencia
Los resultados del presente
estudio, para este parámetro, difieren de los reportados por Alanís-Rodríguez y
col. (2008); Jiménez-Pérez y col. (2012) y Pequeño-Ledezma y col. (2012),
quienes evaluaron áreas del MET con algún grado de disturbio, ya que
registraron no más de tres especies presentes en todos los sitios de muestreo.
Sin embargo, los resultados de este trabajo, concuerdan con los de Mora-Donjuán
y col. (2013a), estudio de un área sin alteración reciente, donde se
registraron que siete especies se encontraban en todos los sitios de muestreo.
Lo anterior, prueba que, conforme más especies se encuentren en todos los
sitios de muestreo, menos disturbio muestra el área evaluada, ya que, al no
haber afectaciones graves en la comunidad vegetal, más especies tienen
posibilidad de establecerse en diversos sitios, evitando que predominen un
número reducido de especies en toda el área.
Índice
de valor de importancia (IVI)
En el presente estudio, Prosopis leavigata fue
la especie con mayor IVI (28.59 %),
con una diferencia porcentual de 15.18 %, respecto a la siguiente especie Havardia pallens. Este
estudio es el segundo que registra a Prosopis leavigata como la especie con el mayor IVI en el MET con uso ganadero
(Alanís-Rodríguez y col., 2008; Jiménez-Pérez y col., 2012; Pequeño-Ledezma y
col., 2012; Mora-Donjuán y col., 2013a; 2013b).
Prosopis leavigata presentó una densidad relativa (20.39 %) ligeramente
superior a la de Havardia pallens
(19.09 %). Sin embargo, su dominancia relativa (56.09) fue mucho mayor, con una
diferencia de 44.24 % (Tabla 2). Esto indica una alta preferencia del manejador
para conservar dicha especie, ya que, si se compara con otros estudios, que se
han llevado a cabo para el MET, se observa que son otras las especies con mayor
densidad, entre ellas, Diospyros texana y las del género Vachellia
(Alanís-Rodríguez y col., 2008; Pequeño-Ledezma y col., 2012; Jiménez-Pérez y
col., 2013; Mora-Donjuán y col., 2013a). Esta preferencia se adjudica al
conocimiento sobre los múltiples usos del Prosopis leavigata (mejor conocido como mezquite)
en la región, tales como leña, fuente de carbón, uso artesanal y de forraje
(Estrada-Castillón y col., 2005), siendo el último de especial interés para la
producción ganadera (Ríos-Saucedo y col., 2012).
Distribución
vertical de las especies
Aunado a los resultados y
siendo que el diámetro menor registrado fue de 4 cm (datos no mostrados), se
tiene una comunidad vegetal que no se encontraba en regeneración, como otros
matorrales (Alanís-Rodríguez y col., 2011; Jiménez-Pérez y col., 2012; Mora-Donjuán
y col., 2013b), pero que tampoco tiene las condiciones óptimas de un matorral
primario (Mora-Donjuán y col., 2013b; Yerena-Yamallel
y col., 2014); sino que sus componentes conforman una masa irregular joven e incoetánea, en la cual, los árboles crecen principalmente
en diámetro (Araujo e Iturre, 2006).
Diversidad
El valor de DMg, registrado
en este estudio, fue superior al evaluado por Alanís-Rodríguez y col. (2008),
en un área con historial de uso de ganadería intensiva y extensiva. También el
índice de diversidad de Shannon-Wiener (H´), reportado en este trabajo, fue
superior a los obtenidos en áreas con diferente historial de uso productivo
(Alanís-Rodríguez y col., 2008; Jiménez-Pérez y col., 2012; Pequeño-Ledezma y
col., 2012; Mora-Donjuán y col., 2013b; Molina-Guerra y col., 2013) e incluso
comparado con áreas de MET sin alteración reciente (Mora-Donjuán y col., 2013a;
2013b). Una explicación de que los valores reportados en el estudio fueron
superiores, con respecto a los reportados en las investigaciones mencionadas,
sería que la comunidad evaluada se encontraba en un estado joven, donde la
constante selección de especies crea nuevos espacios, que permiten que se
establezcan otras especies o nuevos individuos.
CONCLUSIONES
La composición de especies
arbóreas y arbustivas en el área de estudio conserva atributos cercanos a las
comunidades de matorral espinoso tamaulipeco (MET) sin alteraciones, como la
riqueza de especies, diversidad, densidad y área de copa. Sin embargo, en el
manejo de selección de las especies destaca la preferencia por conservar
individuos de altura distribuidas en el estrato II (6.16 m a 9.84 m), con
diámetros de 10-20 cm, características de una comunidad vegetal irregular e incoetánea. La especie de preferencia fue Prosopis laevigata,
deseable por ser multipropósito (fuente de leña y carbón, uso artesanal y
forrajera). Se considera importante aumentar la investigación de los
aprovechamientos mixtos en el noreste de México.
AGRADECIMIENTOS
Al Programa de Mejoramiento del Profesorado
(PROMEP) por el financiamiento de la investigación mediante el proyecto
“Análisis de la regeneración natural del matorral espinoso tamaulipeco en el
noreste de México (PROMEP/ 103.5/12/3585)”. A Guadalupe Pérez, Mario Moreno y
Juan Garza por su colaboración en el trabajo de campo.
REFERENCIAS
Alanís-Flores,
G. J. (1996). Vegetación y flora de Nuevo
León, una guía botánico-ecológica. San Nicolás de los Garza: Impresora
Monterrey, S.A. de C.V. 251 Pp.
Alanís-Rodríguez,
E., Jiménez-Pérez, J., Aguirre-Calderón, O. A., Treviño-Garza, E. J.,
Jurado-Ybarra, E. y González-Tagle, M. A. (2008). Efecto del uso del suelo en
la fitodiversidad del matorral espinoso tamaulipeco. Ciencia UANL. 11(1): 56-62.
Alanís-Rodríguez, E., Jiménez-Pérez, J., Canizales-Velázquez, P. A., González-Rodríguez, H.,
Mora-Olivo, A. (2015). Estado actual del conocimiento de la estructura arbórea
y arbustiva del matorral espinoso tamaulipeco del noreste de México. Revista Iberoamericana de Ciencias.
2(7): 69-80.
Alanís-Rodríguez,
E., Jiménez-Pérez, J., González-Tagle, M. A., Yerena-Yamallel,
J. I., Cuellar-Rodríguez, L. G. y Mora-Olivo, A. (2013). Análisis de la
vegetación secundaria del matorral espinoso tamaulipeco, México. Phyton. 82(2):
185-191.
Alanís-Rodríguez, E., Jiménez-Pérez, J., ValdecantosDema, A., Pando-Moreno, M., Aguirre-Calderón, O.
y Treviño-Garza, E. J. (2011). Caracterización de regeneración leñosa postincendio de un ecosistema templado del Parque Ecológico
Chipinque, México. Revista Chapingo Serie Ciencias Forestales y del Ambiente. 17(1):
31-39.
Araujo, P. A. e Iturre M. C.
(2006). Ordenación de Bosques
Irregulares. Cátedra de Ordenación Forestal. Facultad de Ciencias
Forestales, Universidad Nacional de Santiago del Estero. [En línea]. Disponible
en: http://fcf.unse.
edu.ar/archivos/series-didac-ticas/SD-28-Ordenacionbosques-irregulares-ARAUJO.pdf.
Fecha de consulta: 20 de noviembre de 2013.
Arriaga-Cabrera, L. (2009). Implicaciones del cambio de
uso de suelo en la biodiversidad de los matorrales xerófilos: un enfoque multiescalar. Investigación
ambiental. 1(1): 6-16.
Barrance, A., Schreckenberg, K., and Gordon J. (2009). Conservation through use: lessons from the
Mesoamerican dry forest. Londres: Overseas Development Institute. 124 Pp.
Cantú-Silva, I., Estrada-Castillón, E.,
González-Rodríguez, H. y Delgado-Villalobos, J. A. (2017). Estructura y
diversidad del matorral desértico rosetófilo
rehabilitado con rodillo aireador, Coahuila, México. Polibotánica. (44): 95-107.
Clifford, H. y Stephenson W. (1975). An Introduction to Numerical
Classification. Nueva York: Academic
Press. 229 Pp.
CONABIO, Comisión Nacional para el conocimiento y Uso de
la Biodiversidad (2009). Estado de
Conservación y Tendencias de Cambio, en Capital Natural de México, vol. 2.
[En línea]. Disponible en: http://www.biodiver sidad.gob.mx/pais/pdf/CapNatMex/Vol%20II/II00_Preli
minares%20Guia%20de%20lectura%20Indice%20Siglas. pdf. Fecha de consulta: 1 de noviembre de 2013.
CONAFOR,
Comisión Nacional para el conocimiento y Uso de la Biodiversidad (2010). Paquete tecnológico: Sistemas silvopastoriles. [En línea]. Disponible en: http://
www. conafor.gob.mx/biblioteca/SISTEMAS-SILVOPASTORIL.pdf. Fecha de consulta:
13 de diciembre de 2013.
Cram, S., Sommer, I., Morales, L. M., Oropeza, O., Carmona, E., and
González-Medrano, F. (2006). Suitability of the vegetation types in Mexico’s Tamaulipas state for the
siting of hazardous waste treatment plants. Journal of Environmental Management. 80(1): 13-24.
Estrada-Castillón,
E., Villarreal-Quintanilla, J. y Jurado, E. (2005). Leguminosas del norte del
estado de Nuevo León, México. Acta Botanica Mexicana. (73):1-18.
FAO, Food and Agriculture Organization of the United Nations (2016). Global Forest Resources Assessment 2015, How are the world’s
forests changing? (Second edition). Roma: Food and Agriculture Organization of the United Nations. 44 Pp.
Foroughbakhch, R.,
Hernández-Piñero, J. L., Alvarado Vásquez, M. A., Céspedes-Cabriales,
E., Rocha-Estrada, A., and Cárdenas-Ávila, M. L. (2009). Leaf
biomassdetermination on woody shrub species
in semiarid zones. Agroforestry Systems.
77(3): 181-192.
García-Hernández.
J. y Jurado, E. (2008). Caracterización del matorral con condiciones prístinas
en Linares, N. L., México. Ra Ximhai. 4(1): 1-21.
González-Rodríguez,
H., Ramírez-Lozano, R. G., Cantú-Silva, I., Gómez-Meza, M. V. y Uvalle-Sauceda, J. I. (2010). Composición y estructura de
la vegetación en tres sitios del estado de Nuevo León, México. Polibotánica.
(29): 91-106.
INEGI, Instituto
Nacional de Estadística y Geografía (1986). Síntesis Geográfica de Nuevo León.
[En línea]. Disponible en: http://www.beta.inegi.org.mx/app/biblioteca/ ficha.html?upc=702825220747. Fecha de consulta: 1 de
noviembre de 2013.
INEGI, Instituto
Nacional de Estadística y Geografía (2013). Conjunto de datos vectoriales de
uso de suelo y vegetación, escala 1:250 000, serie V. [En línea]. Disponible
en:
http://catalogo.datos.gob.mx/dataset/mapas-de-usodel-suelo-y-vegetacion-escala-1-250-000-serie-v-nuevoleon.
Fecha de consulta: 1 de noviembre de 2013.
INEGI, Instituto
Nacional de Estadística y Geografía (2017). Encuesta Nacional Agropecuaria ENA
2017: Conociendo el campo de México, Resultados. [En línea]. Disponible en:
http://www.beta.inegi.org.mx/contenidos/pro gramas/ena/2017/doc/ena2017_pres.pdf. Fecha de consulta: 28 de febrero de
2019.
Jiménez-Pérez,
J., Alanís-Rodríguez, E., González-Tagle, M. A., Aguirre-Calderón, O. A., and
Treviño-Garza, E. J. (2013). Characterizing regeneration of woody species in areas with different land history tenure
in the tamaulipan thornscrub, Mexico. The Southwestern Naturalist. 58(3): 299-304.
Jiménez-Pérez,
J., Alanís-Rodríguez, E. A., Ruíz-González, J. L., González-Tagle, M. A., Yerena-Yamallel, J. I. y Alanís-Flores, G. J. (2012).
Diversidad de la regeneración leñosa del matorral espinoso tamaulipeco con
historial agrícola en el noreste de México. Ciencia
UANL. 15(58): 66-71.
Jose, S. (2012). Agroforestry for conserving and enhancing biodiversity. Agroforestry Systems. 85(1): 1-8.
Maass, J. M.,
Balvanera, P., Castillo, A., Daily, G. C., Mooney, H. A., Ehrlich, P., ..., and Martínez-Yrizar, A.
(2005). Ecosystem services
of tropical dry forests: insights from long-term
ecological and social research
on the Pacific
Coast of Mexico. Ecology and Society.
10(1): 1-23.
Magurran, A. (2004). Measuring Biological Diversity. Oxford: Blackwell
Publishing Company. 264 Pp.
Mahecha, L. y Zoot, M. S. (2002). El silvopastoreo:
una alternativa de producción que disminuye el impacto ambiental de la
ganadería bovina. Revista Colombiana de
Ciencias Pecuarias. 15(2): 226-231.
Martella, M., Trumper, E., Bellis, L., Renison, D., Giordano, P., Bazzano,
G. y Raquel, M. (2012). Manual de Ecología: Evaluación de la biodiversidad. Reduca (Biología) Serie Ecología. 5(1):
71-115.
Mitlohner, F. M., Galyean, M. L., and McGlone, J.
J. (2002). Shade effects on performance, carcass traits, physiology, and behavior of heat-stressed feedlot heifers. Journal of Animal Science.
80(8): 2043-2050.
Molina-Guerra, V. M., Pando-Moreno, M., Alanís-Rodríguez,
E., Canizales-Velázquez, P. A., González-Rodríguez,
H. y Jiménez-Pérez, J. (2013). Composición y diversidad vegetal de dos sistemas
de pastoreo en el matorral espinoso tamaulipeco del Noreste de México. Revista Mexicana de Ciencias Pecuarias.
4(3): 361-371.
Mora-Donjuán, C. A., Alanís-Rodríguez, E., JiménezPérez, J., González-Tagle, M. A., YerenaYamallel, J. I. y Cuellar-Rodríguez, L. G. (2013a).
Estructura, composición florística y diversidad del matorral espinoso
tamaulipeco, México. Ecología Aplicada.
12(1): 29-34.
Mora-Donjuán, C.
A., Jiménez-Pérez, J., Alanís-Rodríguez, E., Rubio-Camacho, E., Yerena-Yamallel, J. I. y González-Tagle, M. A. (2013b).
Efecto de la ganadería en la composición y diversidad arboréa
y arbustiva del matorral espinoso tamaulipeco. Revista Mexicana de Ciencias Forestales. 4(17): 124-137.
Moreno-Medina,
S., Ibarra-Flores, F. A., Martín-Rivera, M. H., Moreno-Álvarez, C. y
Retes-López, R. (2017). Impacto económico del deterioro de los recursos
naturales asociados con la eficiencia de la reproducción ganadera. Revista Mexicana de Agronegocios.
21(40): 649-659.
Müeller-Dombois, D. and Ellenberg, H. (1974). Aims and methods of vegetation
ecology. Nueva York: John Wiley
& Sons Inc. 547 Pp.
Musálem, M. A. (2002). Sistemas
agrosilvopastoriles: Una alternativa de desarrollo
rural sustentables para el trópico mexicano. Revista Chapingo Serie Ciencias Forestales y del Ambiente. 8(2):
91-100.
Návar-Chaidez, J. (2008). Carbon fluxes resulting
from land-use changes in the Tamaulipan thornscrub of northeastern Mexico. Carbon Balance Management. 3(1): 6.
Pando-Moreno, M. and
Villalón-Mendoza, H. (2001). Potential agroforestry species identified in the Tamaulipan thornscrub of north-eastern Mexico. Agroforestry Today.
13(1-2): 14-15.
Pequeño-Ledezma, M. A., Alanís-Rodríguez, E.,
Jiménez-Pérez, J., Aguirre-Calderón, O. A., González-Tagle, M. A. y
Molina-Guerra, V. M. (2017). Análisis estructural de dos áreas del matorral
espinoso tamaulipeco del noreste de México. Madera
y Bosques. 23(1): 121-132.
Pequeño-Ledezma,
M. A., Alanís-Rodríguez, E., Jiménez-Pérez, J., González-Tagle, M. A., Yerena-Yamallel, J. I., Cuellar-Rodríguez, G. y Mora-Olivo,
A. (2012). Análisis de la restauración pasiva postpecuaria
en el matorral espinoso tamaulipeco del noreste de México. CienciaUAT. 24(2):
42-47.
Pretzsch, H. (1996). Strukturvielfalt als Ergerbnis Waldbaulichen Handels. Nehresheim: Deutscher Verband Forstlicher Forschungsanstalten. Sekt. Ertragskunde. 849 Pp.
Ramírez-Marcial,
N., Rueda-Pérez, M. L., Ferguson, B. G. y
Jiménez-Ferrer, G. (2012). Caracterización del sistema agrosilvopastoril
en la Depresión Central de Chiapas. Avances
de Investigación Agropecuaria. 16(2): 7-22.
Ríos-Saucedo, J.
C., Valenzuela-Nuñez, L. M., RiveraGonzález,
M., Trucíos-Caciano, R. y Sosa-Pérez, G. (2012).
Diseño de un sistema sil-vopastoril en zonas
degradadas con mezquite en Chihuahua, México. Tecnociencia Chihuahua. 6(3): 174-180.
Rodríguez-Estrella,
R., Benítez-Malvido, J., Del-ValDe-Gortari, E. y
Santos-Barrera, G. (2016). Impacto de las actividades humanas en la
biodiversidad y en los ecosistemas. En P. Balvanera, E. Arias, R. RodríguezEstrella, L. Almeida-Leñero, J. J. Schmitter-Soto (Eds.), Una
mirada al conocimiento de los ecosistemas de México (pp. 57-189). Ciudad de
México, México: Universidad Nacional Autónoma de México.
Rosales-Adame,
J. J., Cuevas-Guzmán, R., Gliessman, S. R., and Benz,
B. F. (2014). Estructura y diversidad arbórea en el Sistema agroforestal de
piña bajo sombra en el Occidente de México. Tropical
and Subtropical Agroecosystems. 17(1): 1-18.
Rosete-Vergés, F. A., Pérez-Damián, J. L., VillalobosDelgado,
M., Navarro-Salas, E. N., Salinas-Chávez, E. y Remond-Noa,
R. (2014). El avance de la deforestación en México 1976-2007. Madera y Bosques. 20(1): 21-35.
Rubio-Camacho,
E. A., González-Tagle, M. A., Jiménez-Pérez, J., Alanís-Rodríguez, E. y
Ávila-Flores, D. (2014). Diversidad y distribución vertical de especies
vegetales mediante el índice de Pretzsch. Ciencia UANL. 17(65): 34-41.
SAGARPA, Secretaría de Agricultura, Ganadería, Desarrollo
Rural, Pesca y Alimentación (2013). Sistemas Silvopastoriles.
México: Subsecretaría de desarrollo rural, SAGARPA. [En línea]. Disponible en:
http://www.sagarpa. gob.mx /desarrolloRural /Documents /fichasaapt /
Sistemas%20silvopastoriles.pdf. Fecha de consulta: 15 de diciembre de 2013.
Sánchez, M. D.,
Rosales, M. y Murgueitio E. (2003). Agroforestería en América Latina. En M. D. Sánchez y D.
Rosales-Méndez (Eds.), Agroforestería para la producción animal en América Latina
II (pp. 1-12). Roma, Italia: FAO.
Santos, T., y Tallería, J. L. (2006). Pérdida y fragmentación del
hábitat: efecto sobre la conservación de las especies. Ecosistemas. 15(2): 3-12.
Sarukhán, J., Urquiza-Haas, T., Koleff, P., Carabias, J., Dirzo, R., Ezcurra,
E., and Soberón, J. (2015). Strategic
Actions to Value, Conserve,
and Restore the Natural
Capital of Megadiversity Countries:
The Case of Mexico. BioScience. 65(2): 164-173.
Shannon, C. E.
and Weaver, W. (1949). The Mathematical Theory
of Communication. Illinois, EUA: University of Illinois Press. 144
Pp.
Williams-Linera,
G. (2002). Tree species richness complementarity, disturbance and fragmentation in
a Mexican tropical montane cloud forest. Biology and Conservation. 11(10): 1825-1843.
Yerena-Yamallel, J. I.,
Jiménez-Pérez, J., Alanís-Rodríguez, E., Aguirre-Calderon,
O. A., González-Tagle, M. A., and Treviño-Garza, E. J. (2014). Dinámica de la
captura de carbono en pastizales abandonados del noreste de México. Tropical and Subtropical Agroecosystems.
17: 113-121.