Efecto de Candida norvegensis sobre la degradabilidad ruminal de rastrojo de
maíz y el crecimiento de corderos
Effect
of Candida norvegensis on ruminal
degradation of cornstover and on growth performance of lambs
Acción
de levaduras en la degradacion de fibra
Oscar Ruiz-Barrera1, Jesús Lopéz-Morones1,
Jaime Salinas-Chavira2, Yamicela Castillo-Castillo1*
*Correspondencia:
ycastillo75@yahoo.com/Fecha
de recepción: 7 de diciembre de 2018/Fecha
de aceptación: 25 de septiembre de 2019/Fecha
de publicación: 31 de enero de 2020
1Universidad
Autónoma de Chihuahua, Facultad de Zootecnia y Ecología. Periférico Francisco
R. Almada km 1, col. Zootecnia, Chihuahua, Chihuahua, México, C. P. 31453. 2Universidad
Autónoma de Tamaulipas, Facultad de Medicina Veterinaria y Zootecnia, Ciudad
Victoria, Tamaulipas, México.
Resumen
Los probióticos favorecen el desarrollo de
microorganismos benéficos en el rumen, lo que
incrementa la digestibilidad de los
nutrientes y mejora el desempeño productivo de los rumiantes; con esto,
se tiene la posibilidad de utilizar
ingredientes como el rastrojo de maíz de relativo bajo valor nutritivo, pero
altamente disponible en algunos lugares a bajo precio. Convencionalmente,
se utilizan como probióticos las levaduras Saccharomyces, aunque existen
reportes sobre el uso de cepas autóctonas, como Candida norvegensis. El objetivo de este estudio fue evaluar
los efectos del probiótico de Candida norvegensis en la
degradabilidad ruminal in situ de rastrojo
de maíz y en el comportamiento
productivo de ovinos en crecimiento. La levadura Candida norvegensis (cepa
Levazoot 15) [0 g (T1) y 5 g (T2)] se usó para determinar la degradación ruminal in situ (DRMS), del
rastrojo de maíz, en 3 vacas canuladas ruminalmente por medio de la técnica de
la bolsa de poliéster. No hubo efecto de la levadura (P > 0.05) para la
fracción (a), (b) y (a+b); pero la degradabilidad efectiva al 1 %/h y 5 %/h de
recambio ruminal fue mayor para T2 (P < 0.05). En un segundo experimento, 32
corderos se asignaron al azar a corrales individuales por 105 d para evaluar 4
dietas que difirieron en la proporción de
concentrado/forraje: T1 = 75:25, T2 =
75:25, T3 = 50:50, y T4 = 25:75.
A excepción de T1, las dietas fueron suplementadas con Candida norvegensis,
a razón de 15 mL/kg de peso vivo, equivalente a 5 g/d de levadura en base seca.
Los ovinos en la dieta con 75 % de concentrado más la levadura (T2) presentaron
mayor ganancia de peso, y mejor conversión alimenticia (P < 0.05). Se
concluye que Candida norvegensis mostró efectos benéficos en la
degradabilidad ruminal y en el
desarrollo de corderos.
Palabras clave: levaduras; degradabilidad
ruminal; corderos; probiótico.
Abstract
Probiotics assist in the development of
beneficial microorganisms in the rumen that increase digestibility of nutrients and improves the productive performance of ruminants; it also has the possibility of using ingredients as
corn stover of relatively low nutritional
value, but available in some places
at low prices. Saccharomyces yeasts are conventionally used as
probiotics and there are reports that use
native strains such as Candida
norvegensis. The objective of
this study was to evaluate the effects of the probiotic of Candida
norvegensis on the in situ ruminal dry matter degradability of corn stover
and on the productive performance of growing
lambs. In the first experiment, the yeast
Candida norvegensis (strain Levazoot 15) [0 g (T1)] and 5 g (T2) was used to determine the in situ ruminal
dry matter degradation (RDMD) of the corn stover in 3 cows with cannulas
in the rumen, which was determined by the polyester bag technique. There was
no effect of yeast (P > 0.05) on fraction (a), (b) and (a+b). However, the effective degradability at 1 %/h and 5
%/h of ruminal turnover was higher in T2 (P < 0.05). In the second experiment,
32 lambs were randomly assigned to
individual pens for 105 d to evaluate 4 diets that differed in the
proportion of concentrate: forage: T1 = 75:
25, T2 = 75:25; T3 = 50: 50, and T4 = 25: 75. With the exception of T1,
the diets were supplemented with Candida
norvegensis at 15 mL/kg of live weight, equivalent to 5 g/d of yeast in dry matter basis. The lambs in the diet with
75 % of concentrate plus the yeast (T2)
showed greater weight gain and best feed
conversion (P < 0.05). It is concluded that Candida norvegensis
showed beneficial effects on ruminal degradability and on the growth of
lambs.
Keywords: yeast; ruminal
degradability; lambs; probiotic.
Introducción
La función digestiva de los rumiantes es
fundamental en las variables de comportamiento de la producción ganadera
(Dittmann y col., 2015). De acuerdo con Kannan y col. (2014), las continuas prohibiciones de aditivos, como
los antibióticos, el costo elevado de insumos (Leibtag, 2008), y la escasez de
zonas de pastoreo (Tamburello y col., 2014), han promovido la búsqueda de
opciones para disminuir los costos de producción, sin afectar la salud de
animales y de humanos. Una alternativa son los probióticos, los cuales poseen
la capacidad de mejorar la función digestiva de los animales, contribuyendo a
reducir la inversión por concepto de alimentación de las explotaciones de
ovinos, caprinos y bovinos, mediante la utilización de ingredientes económicos,
pero que carecen de un valor nutritivo adecuado. Por lo tanto, una mejora en la
funcionalidad del tracto digestivo de los rumiantes, que favorezca el ambiente
ruminal para el desarrollo de microorganismos
benéficos, como las bacterias y hongos celulolíticos,
podría transformarse en beneficios posteriores, como una mejor
conversión alimenticia y mayor ganancia diaria de peso, que se reflejaría en
importantes ahorros de índole alimenticia y un uso más intensivo de esquilmos
agrícolas (Salinas-Chavira y col., 2013; Abdel-Aziz y col., 2015; Gadekar y
col., 2015; Puniya y col., 2015). Sin embargo,
existen pocos estudios en relación al uso de cepas de levaduras
diferentes a las de Saccharomyces, las cuales, en algunos casos,
promueven mejoras ruminales y hacen más eficiente la producción animal, siendo
prioritario profundizar en los estudios de la utilización
de cepas autóctonas. Candida norvegensis (cepa Levazoot
15) ha demostrado, en estudios in vitro, tener efectos positivos para
favorecer el desarrollo de la flora nativa y ser capaz de desdoblar incluso las
fracciones menos digestibles de la fibra (Castillo-Castillo y col., 2016; Ruiz
y col., 2016). El presente estudio es relevante, dado que existe poca
información de levaduras no-Saccharomyces, de cepas autóctonas aisladas en la
zona de estudio, que puedan actuar como probióticos, y que permitan mejorar el
crecimiento de ovinos alimentados con
rastrojo de maíz, ingrediente alimenticio comúnmente usado en raciones para la engorda de corderos en México.
El objetivo del presente trabajo fue
evaluar los efectos probióticos de Candida norvegensis en la
degradabilidad ruminal in situ de rastrojo de maíz, y sobre el
comportamiento productivo de ovinos
en crecimiento.
MATERIALES Y MÉTODOS
Lugar de trabajo
El presente estudio se llevó a cabo en la
Facultad de Zootecnia y Ecología de la Universidad
Autónoma de Chihuahua, localizada geográficamente a 28°38’ latitud N y 106°04’
longitud W, a una altitud de 1 435 msnm, con una precipitación media
anual de 387.5 mm y una temperatura media anual de 18.2 °C de acuerdo al
Instituto Nacional de Estadística y Geografía (INEGI, 2016). Todos los animales
utilizados en este estudio fueron manejados de acuerdo al Comité de Bioética de la Universidad Autónoma de Chihuahua. El
estudio se realizó del 25 de marzo al 15 de junio de 2015.
Experimento de
degradabilidad ruminal en ganado
bovino
Animales y su manejo
Para evaluar la degradabilidad ruminal del
rastrojo de maíz se utilizaron 3 vacas adultas, con peso promedio de 496 ± 1.5
kg, equipadas con cánula ruminal permanente, suplementadas con un aditivo (probiótico)
de levaduras Candida norvegensis (cepa Levazoot 15). Previamente, las
vacas fueron vacunadas para prevenir contra enfermedades infecciosas respiratorias,
diarreas virales y leptospirosis. Además, fueron desparasitadas con Ivermectina
e inyectadas vía intramuscular con suplemento de selenio y vitamina E. Las
vacas fueron alojadas individualmente y alimentadas ad libitum con una
dieta basal de alfalfa y rastrojo de maíz (50:50), la cual satisface los requerimientos de nutrientes para mantenimiento
de las vacas, y permite el crecimiento de
bacterias fibrolíticas en rumen. El alimento fue suministrado
diariamente a las 10:00 horas y 17:00 horas, y el agua fue administrada a libre
acceso.
Tratamientos
experimentales y manejo de la levadura
Se evaluaron 2 tratamientos
usando 2 periodos experimentales, en los cuales, los
animales fueron alimentados con la dieta
basal. En el primer periodo las vacas no recibieron el probiótico (T1:
control). En el segundo, se suplementaron diariamente, vía ruminal, con 15 mL/kg
de peso vivo de probiótico líquido a base de la levadura Candida norvegensis
(cepa Levazoot 15), equivalente a 5 g en base seca (T2).
Para la preparación del inóculo de
levadura, se tomó una muestra del medio de cultivo de la cepa Levazoot 15 con
24 h de crecimiento, y se disolvió en 10 mL de caldo extracto de malta (DIFCO
St Louis MO, USA); se incubó en un tubo de ensayo de 100 mL a 30 °C por 24 h.
Se mezclaron 50 mL de este cultivo con 50 mL de caldo extracto de malta en un
matraz estéril Erlenmeyer de 100 mL. La mezcla nuevamente se incubó como se
mencionó previamente (Castillo-Castillo y col., 2016). El cultivo resultante
fue usado como inóculo para escalar la producción de la cepa Levazoot 15 en un
fermentador dinámico, con capacidad de producción de 30 L, con una fase de
descanso de 2 h y otra de actividad
de 15 min.
Degradabilidad
ruminal in situ de las dietas
Para la determinación de la degradabilidad
ruminal in situ, de la materia seca, se consideraron dos periodos
experimentales consecutivos (el periodo 1 de 30 d, y el periodo 2 de 31 d). En
el primer periodo las vacas recibieron la dieta base sin levadura, y en el segundo
periodo, los animales recibieron la dieta base más el aditivo (levadura). En cada
periodo se usaron los mismos animales y se consideraron los últimos 3 d de cada
periodo para la incubación ruminal de
rastrojo de maíz; el resto de los días, en cada periodo, fue de
adaptación. La incubación se realizó de acuerdo a los procedimientos descritos
por Noziére y Michaelt-Doreau (2000), para
lo cual se utilizaron bolsas ANKOM (Macedon, NY. USA), de poliéster,
libres de nitrógeno, con dimensiones de 10
cm x 20 cm y un tamaño de poro de 50 μ (± 15). La muestra de rastrojo de maíz se
secó previamente (55 °C, 72 h) y se molió en un molinoWiley No. 4, equipado con una malla de 2 mm. Se colocaron 3 g de
muestra en cada bolsa. Las bolsas fueron colocadas en el rumen por triplicado
por cada tiempo de incubación, los cuales fueron de 6 h, 12 h, 24 h, 48 h y 72
h. Después de cada tiempo de incubación, las bolsas se lavaron con agua de la llave, hasta que el agua de lavado se tornó
cristalina, y se secaron en una estufa de aire forzado (Techno ® modelo 17;
Chicago, lL, USA) a 60 °C durante 48 h.
La degradabilidad in situ de la MS,
para las muestras, en cada tiempo de incubación, fue calculada como la pérdida de peso de las muestras en las bolsas antes y
después de la incubación ruminal, de acuerdo con el modelo de Ørskov y McDonald (1979), modificado
por McDonald (1981):
![]()
Donde:
P es la desaparición ruminal de la MS al
tiempo t; a es la fracción rápidamente soluble; b es la fracción degradable; c
es la tasa fraccional de degradación por hora de b; a + b es la degradabilidad
potencial (%), la cual representa la cantidad de muestra que puede ser
solubilizada o degradada en el rumen, si el tiempo no es un factor limitante; t
es el tiempo de incubación en el rumen en horas. La fase “lag” es el tiempo (horas)
de la colonización microbiana del sustrato en rumen.
Las constantes de tasa de pasaje ruminal
(k) a 1 %/h y 5 %/h fueron usadas para calcular la degradación efectiva (DE) de la MS de acuerdo al modelo de
Ørskov y McDonald (1979):
DE = a + b (b x c) / (c + k)
Análisis estadístico
Se utilizó un diseño experimental completamente
al azar en donde se consideró como efecto fijo al tratamiento y como efectos
aleatorios a los animales y a los periodos. Los datos fueron analizados con
PROC MIXED de SAS. Las medias se compararon con la prueba de t de
Student con el Software de Análisis Estadístico (SAS, por sus siglas en inglés: Statistitical Analysis System) (SAS, 2007).
Los valores de P < 0.05 fueron considerados estadísticamente significativos.
Experimento de
comportamiento productivo en ovinos
Ovinos y su manejo
Para la prueba de crecimiento se
utilizaron 32 corderos de pelo de cruza pelibuey, dorper y katahdin, con un
peso promedio inicial de 18.14 kg ± 0.25 kg, los cuales fueron agrupados por
peso en 4 bloques y asignados al azar a
corraletas individuales (8 corraletas/tratamiento) de 2.5 m x 2.5 m con
techado de lámina, comedero y bebedero
automático. Los corderos fueron vacunados para prevenir contra
enfermedades infecciosas respiratorias, diarreas virales y leptospirosis;
además, fueron desparasitados con Ivermectina e inyectados vía intramuscular
con suplemento de selenio y vitamina E. Los animales fueron identificados con
aretes al inicio del experimento. Se midió la ganancia diaria de peso (GDP), mediante
un pesaje inicial y sub-secuentemente cada 15
d, en una báscula electrónica (True-Test®, EziWeigh; Columbus, OH, USA)
con capacidad de 200 kg y aproximación de 0.025 kg. El alimento se ofreció diariamente
a las 08:00 horas, y el consumo de agua fue ad libitum. En cada etapa se estimó el consumo de materia seca (CMS)
con base en el alimento ofrecido menos el rechazado. El alimento ofrecido y el
rechazado fueron pesados diariamente, y el nivel de alimentación diaria ofrecida fue 10 % más del consumo de alimento del día anterior. La conversión
alimenticia (CA) se estimó con base en el alimento consumido dividida entre la
ganancia de peso (consumo/ganancia). La fase experimental en la prueba de comportamiento productivo de los ovinos
duró 105 d.
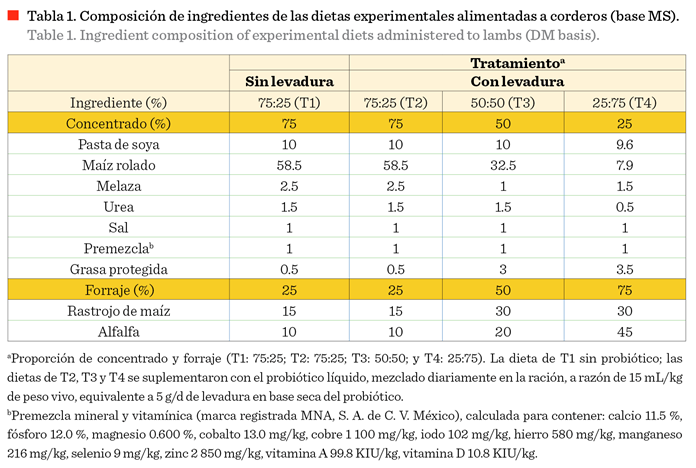
Dietas experimentales
Se probaron 4 dietas diferentes que
variaron en cuanto a la proporción de concentrado y forraje suministrado (T1:
75:25; T2: 75:25, T3: 50:50; y T4: 25:75). Los ingredientes que se incluyeron en el concentrado fueron maíz rolado, pasta de soya, melaza, urea, sal
común, premezcla de minerales para ovinos, y grasa
protegida. Los forrajes considerados en
las dietas fueron rastrojo de maíz y heno de alfalfa. La proporción de
ingredientes de cada dieta se muestra en la Tabla 1. Todos los ingredientes considerados en las dietas se compraron
con un proveedor local dedicado a la venta de alimentos para animales. Las dietas fueron preparadas en el taller de
alimentos balanceados de la Facultad de Zootecnia y Ecología, de la Universidad
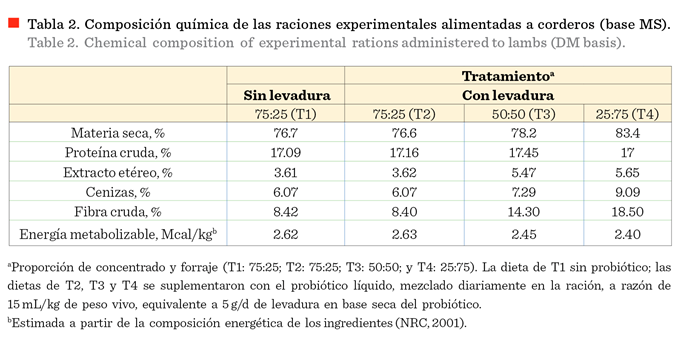
Autónoma de Chihuahua. Las dietas fueron balanceadas con similar contenido de
proteína, pero el nivel de energía disminuyó con el incremento en la proporción de forraje en la dieta (Tabla 2), cuya
composición química fue determinada de acuerdo a la Asociación de Químicos Analíticos Oficiales
(AOAC, por sus siglas en inglés: Association Official Analytical Chemists)
(AOAC, 2005). Todos los corderos con excepción del T1 fueron suplementados con
el probiótico líquido, mezclado diariamente en la ración, a razón de 15 mL/kg
de peso vivo, equivalente a 5 g/d de levadura
en base seca del probiótico en
estudio. La dosis se estimó en base a estudios previos (Ruiz y col.,
2016), y de esta forma mantener la actividad probiótica de la levadura.
Análisis estadístico
Los datos se analizaron con PROC MIXED de
SAS. Se utilizó el peso como covariable continua y una vez ajustados, los
animales fueron asignados al azar a los tratamientos. Las variables de
respuesta fueron: CMS, GDP y CA. Los valores de P < 0.05 fueron considerados
estadísticamente significativos. Las medias de tratamientos se compararon con
la prueba Diferencia Mínima Significativa (SAS, 2007).
RESULTADOS
Experimento de
degradabilidad ruminal en ganado bovino
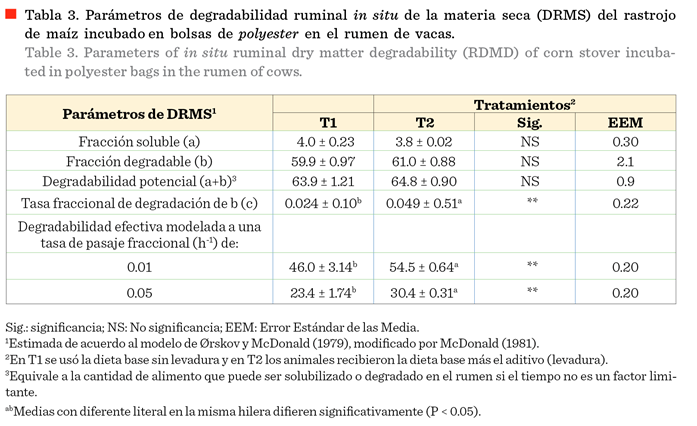
La fracción soluble (a), la fracción
degradable y la degradabilidad potencial de la MS no mostraron diferencia entre
los 2 tratamientos (Tabla 3; P > 0.05). Al contrario, la tasa de degradación (c) fue mayor para T2 que T1 (P < 0.05). La degradabilidad efectiva, tanto
al 1 %/h como al 5 %/h, de recambio ruminal, fue más alta para T2 que T1 (P
< 0.05). Estos resultados coinciden con
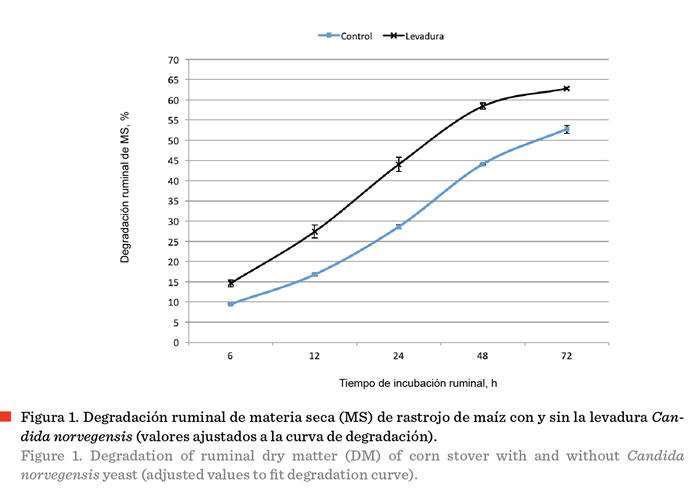
el incremento en la degradación ruminal de MS del rastrojo de maíz, como
consecuencia de la suplementación con la levadura en la dieta (Figura
1).
Comportamiento
productivo en ovinos
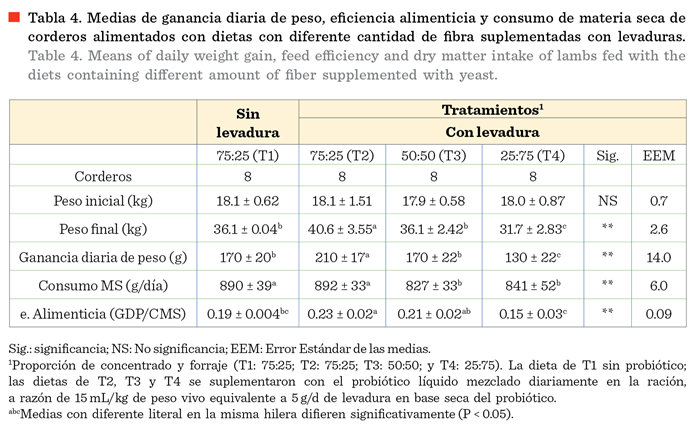
Los resultados se muestran en la Tabla 4.
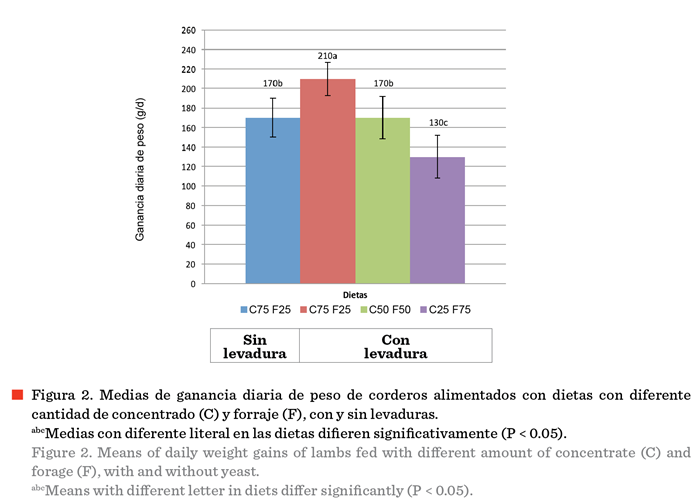
La mayor GDP se observó en los ovinos en la dieta con 75 % concentrado +
levadura (T2); mientras que la menor GDP (P < 0.05) se observó en los ovinos
en la dieta con 25 % de concentrado + levadura (T4). La GDP en los ovinos en la
dieta con 75 % concentrado sin levadura (T1)
y los de la dieta con 50 % de concentrado + levadura (T3) fue similar (P
> 0.05), pero fue menor a los ovinos de T2 y mayor que T4 (P < 0.05). En
la Figura 2 se muestra la GDP obtenida por los ovinos en las dietas
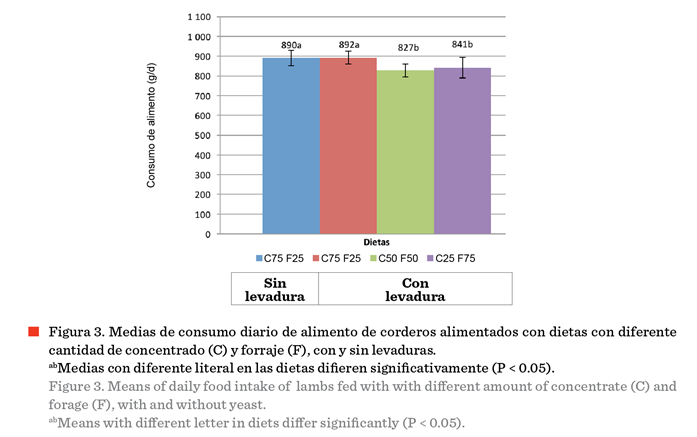
experimentales. Como se observa en la Figura 3, el consumo de MS fue similar en
los ovinos que recibieron las dietas con el
mismo contenido de concentrado sin
(T1) y con (T2) levadura (P > 0.05), y este fue mayor (P < 0.05)
que en los ovinos que recibieron las dietas con menor cantidad de concentrado y levadura (T3 y T4). En la Figura
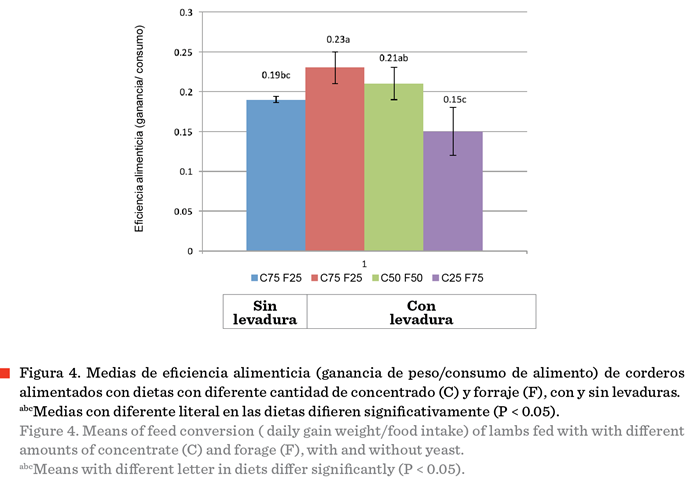
4, se aprecia que la eficiencia alimenticia fue mayor (P < 0.05) en los
ovinos que recibieron la dieta con 75 % de concentrado más levadura (T2), que los ovinos que recibieron la dieta con el mismo
nivel de concentrado pero sin levadura (T1). También se observa que la
eficiencia alimenticia fue similar en los ovinos de T2 y T3 (P > 0.05), y fue menor en los ovinos de T4 (P < 0.05).
DISCUSIÓN
Experimento de
degradabilidad ruminal en ganado bovinos
Los valores de tasa fraccional de
degradación y degradabilidad efectiva del rastrojo de maíz incrementaron cuando
se suplementó la dieta con levadura. En forma consistente, Chaucheyras-Durand y
col. (2015), reportaron mejoras en la degradación de la fracción de fibra
detergente neutro con la adición de Saccharomyces
cerevisiae. Los autores atribuyeron
el efecto de levadura viva sobre la degradación de sustratos fibrosos, por un
aumento en la colonización microbial, adjudicando el efecto no únicamente al pH ruminal, sino también a la utilización
de oxígeno ruminal por las células de levaduras, para una anaerobiosis más eficaz. En forma consistente, Ishaq y col.
(2015) asociaron este efecto a una posible más rápida y efectiva colonización
por microorganismos ruminales. Asimismo, Ruiz y col. (2016) observaron que la
inoculación con Candida norvegensis, a sustratos fibrosos, favorecía el
aprovechamiento de algunas fracciones
altamente indigestibles, por promover
la proliferación de microorganismos celulolíticos, para llevar a cabo
una mejor fermentación ruminal. Promkot y col. (2013), atribuyeron que el uso
de levaduras en rumiantes mejoraba el aprovechamiento de la MS y
su capacidad fermentativa.
Además, Zeoula y col. (2014), reportaron
que la inclusión de probióticos, en las dietas de becerros productores de
carne, ayudó a mejorar la capacidad digestiva de estos, estabilizando el pH en
el rumen, al reducir el lactato y favorecer la proliferación de bacterias
celulolíticas. Castillo-Castillo y col. (2016) y Ruiz y col. (2016), en
estudios in vitro, observaron que la levadura Candida norvegensis disminuyó la concentración de nitrógeno amoniacal,
ácido láctico, así como la producción de gas metano, estabilizando el ambiente
ruminal con valores de pH que rondan el valor neutro, todo lo cual
favorece el desarrollo de microorganismos
degradadores de carbohidratos estructurales. Consistentemente, Bhatt y
col. (2018), en ovinos, detectaron mayor
digestión de la fibra ácido detergente, cuando las dietas se suplementaron con Saccharomyces
cerevisiae. Por su parte, Jia y col. (2018) en dietas para ovinos,
enriquecidas con Bacillus licheniformis y Saccharomyces cerevisiae,
mostraron que la reducción en el amoniaco ruminal se reflejó en mayor síntesis
de proteína microbiana en rumen, lo cual generó mayor retención de nitrógeno en
ovinos, al suplementar dietas para ovinos con Saccharomyces cerevisiae (Obeidat
y col., 2018). En forma diferente, Kowalik y col. (2016), no observaron influencia en la retención de nitrógeno
en ovinos, al suplementar dietas con cultivos vivos de levaduras. Los mismos autores
reportaron que estas diferencias se podrían deber al tipo y dosis de
microorganismos usados, a la diferente composición de ingredientes de las
dietas, así como, a diferencias en los animales usados.
Experimento de comportamiento productivo
en ovinos
En el presente experimento, el
comportamiento productivo de los corderos cambió positivamente por la levadura,
al mejorar las condiciones ruminales y, por ende, aumentando la capacidad de
los microrganismos celulolíticos del rumen, para lograr una mayor tasa
fraccional de degradación ruminal, y de la de gradabilidad efectiva del
rastrojo de maíz, como se observó en el experimento con bovinos. Lo anterior
concuerda con lo reportado por otros autores (Chaucheyras-Durand y col., 2015;
Ishaq y col., 2015). El comportamiento de crecimiento de los corderos se vio
acompañado por consumos adecuados de MS, a pesar de los niveles elevados de
fibra en T3 y T4. Probablemente, debido a un recambio ruminal significativo,
fenómeno que fue reportado en el estudio con bovinos, por influencia de la
levadura en el rumen de los corderos. Estos resultados coinciden con lo obtenido
por Seo y col. (2010), quienes encontraron que las levaduras contribuían significativamente
en la reducción de la presencia de oxígeno, disminuyendo la producción de
lactato, lo que a su vez mejoró la degradabilidad
de los alimentos y la fermentación ruminal,
contribuyendo así, en el desarrollo de
microorganismos celulolíticos, y mejorando por lo tanto la degradabilidad de
las fracciones menos aprovechables de
los alimentos vegetales (Krehbiel y col., 2003; Qadis y col.,
2014; Bernard y col., 2015; Julien y col., 2015; Tristant y Moran, 2015). Otros trabajos, con levaduras no
comerciales, como es el caso del presente estudio, utilizando Candida
norvegensis, demostraron igualmente su eficacia
en la utilización de carbohidratos no estructurales
y el incremento en los parámetros productivos, cuando se inocula el
rumen con Candida tropicalis (Marrero y col., 2014), Candida
pinotopesti (Mokhber-Dezfouli y col.,
2007) y Candida utilis (Ando y col., 2006; Mahyuddin y Winugroho,
2010), mejorando además su condición general.
En este estudio, los
ovinos alimentados en la dieta control (T1; con 25 %
de forraje sin aditivo) mostraron similar ganancia de peso que los ovinos que recibieron la dieta de T3,
en la cual, se incrementó el forraje hasta 50 % de la dieta, lo que representa
un ahorro importante en la alimentación. Esto se debió al efecto benéfico de la
suplementación de levadura; sin embargo, en la dieta de T4 (75 % de forraje más
levadura), los ovinos presentaron la menor eficiencia productiva. Los ovinos que se alimentaron con la dieta
de T2 (25 % de forraje más levadura) tuvieron la mayor ganancia de peso.
En forma consistente, Jia y col. (2018),
en dietas para ovinos en desarrollo (48 % de forraje),
y en finalizado (30 % de forraje), señalaron beneficios en el comportamiento productivo
cuando se suplementó con Bacillus licheniformis y Saccharomyces
cerevisiae. Los autores encontraron aumentos en la fermentación ruminal de
compuestos nitrogenados y en la producción de propionato, además de incremento
en bacterias del género Fibrobacter; también reportaron mejoras en la
capacidad inmune de los ovinos. De acuer-do
con lo anterior, Shankhpal y col. (2016), en dietas con 60 % concentrado
y 40 % forraje, documentaron que, al
suplementar con levaduras vivas de Saccharomyces cerevisae, se mejoró el
comportamiento productivo de cabritos, así como la digestibilidad de nutrientes
en el tubo digestivo total. En contraste, Obeidat (2017), no encontró efecto de
suplementar Saccharomyces cerevisiae en dietas con subproducto de oliva,
en el comportamiento productivo de ovinos en engorda o en la digestibilidad de
nutrientes. El autor señaló que las diferencias en los reportes se pueden deber
en parte a las diferencias en composición de las dietas experimentales, así
como al tipo y a la dosis de levaduras que se emplean en los diferentes estudios.
El consumo de MS por los ovinos, en la dieta
control con 25 % de forraje, sin levadura (T1), fue similar a lo observado en
la dieta con 25 % de forraje con levadura (T2). En este estudio, también se
registró que el consumo de MS fue menor en las dietas con 50 % y 75 % de forraje (T3 y T4), a pesar que
estas dietas contenían Candida norvegensis. Con el mismo nivel de
forraje (T1 vs T2) la levadura no influyó en el consumo de MS. En los estudios
de investigación, las levaduras no muestran efecto constante en el consumo de
MS. De acuerdo con este trabajo, otros autores tampoco reportan efecto de
levaduras en consumo de MS en ovinos (Haddad y Goussous, 2005; Obeidat, 2017;
Jia y col., 2018). Al usar diferente nivel de forraje, en dietas para ovinos,
Issakowicz y col. (2013) no registraron efecto de levadura en consumo de MS. En
forma diferente, Obeidat y col. (2018),
encontraron interacción de forraje con levadura, observando mayor efecto en
consumo de MS en dietas bajas en forraje suplementadas con levadura, mientras
que en dietas altas en forraje la levadura no
tuvo el mismo efecto. El mismo autor también registró menor consumo en dietas
con mayor contenido de forraje.
CONCLUSIONES
De acuerdo con los
resultados de este estudio, el probiótico a base de Candida
norvegensis mostró capacidad de incrementar la degradabilidad ruminal de la
materia seca, además de permitirle a los corderos aprovechar alimentos
con alto contenido de fibra (50 % forraje) e igualar en eficiencia alimenticia
a los
corderos alimentados con dietas ricas en concentrado
(25 % forraje sin levadura), lo que podría ayudar a reducir costos de
alimentación y mantener la eficiencia productiva de los animales. La
suplementación con levadura, en dietas
con 25 % de forraje mejoró en este estudio la ganancia de peso de los ovinos. Se espera realizar una futura
investigación con Candida norvegensis, considerando otras variables de
fermentación ruminal, como pH, ácidos grasos
volátiles, amoniaco, además de hacer conteos de protozoarios.
ReferencIAs
Abdel-Aziz, N. A., Salem, A. Z., El-Adawy, M. M., Camacho,
L. M., Kholif, A. E., Elghandour, M. M., and Borhami, B. E. (2015). Biological
treatments as a mean to improve feed utilization in agricultura animals, an
overview. Journal of Integrative Agriculture. 14(3): 534-543.
Ando, S., Nishiguchi, Y., Hayasaka, K., Iefuji, H., and
Takahashi, J. (2006). Effects of Candida utilis treatment on the
nutrient value of rice bran and the effect of Candida utilis on the
degradation of forages in vitro. Asian-Australasian Journal of Animal Sciences.
19(6): 806-810.
AOAC, Association Official Analytical Chemists (2005). Official
methods of analysis of AOAC International. (18a ed.) Maryland, E. U. A.:
AOAC International. 486 Pp.
Bernard, J. K. (2015). Milk yield and composition of
lactating dairy cows fed diets supplemented with a probiotic extract. The Professional Animal Scientist. 31(4): 354-358.
Bhatt, R. S., Sahoo, A., Karim, S. A., and Gadekar, Y. P.
(2018). Effects of Saccharomyces cerevisiae and rumen bypass-fat
supplementation on growth, nutrient utilisation, rumen fermentation and carcass traits of lambs. Animal Production Science. 58(3): 530-538.
Castillo-Castillo, Y., Ruiz-Barrera, O., Burrola-Barraza,
M. E., Marrero-Rodriguez, Y., Salinas-Chavira,
J., Angulo-Montoya, C., and Camarillo, J. (2016). Isolation and characterization of yeasts from fermented
apple bagasse as additives for ruminant feeding. Brazilian Journal of Microbiology.
47(4): 889-895.
Chaucheyras-Durand, F., Ameilbonne, A., Bichat, A., Mosoni,
P., Ossa, F., and Forano, E. (2015). Live yeasts enhance fibre degradation in
the cow rumen through an increase in plant substrate colonisation by fibrolytic bacteria and fungi. Journal of Applied Microbiology. 120(3): 560-570.
Dittmann, M. T., Hummel, J., Hammer, S., Arif, A., Hebel,
C., Muller, D. W. H., and Clauss, M. (2015). Digesta kinetics in gazelles in
comparison to other ruminants: Evidence for taxon-specific rumen fluid
throughput to adjust digesta washing to the natural diet. Comparative
Biochemistry and Physiology A-Molecular and Integrative Physiology. 185(1):
58-68.
Gadekar, Y. P., Shinde, A. K., Sahoo, A., and Karim, S. A.
(2015). Effect of probiotic supplementation on carcass traits and meat quality
of Malpura lambs. The Indian Journal of Small Ruminants. 21(3): 306-310.
Haddad, S. G. and Goussous, S. N. (2005). Effect of yeast
culture supplementation on nutrient intake, digestibility and growth
performance of Awassi lambs. Animal
Feed Science and Technology. 118(3-4):
343-348.
INEGI, Instituto Nacional de Estadística y Geografía
(2016). Sistema para la Consulta del Anuario Estadístico del Estado de
Chihuahua. [En línea]. Disponible en:
http://cuentame.inegi.org.mx/monografias/informacion/chih/territorio/clima.aspx?tema=me&e=08.
Fecha de consulta: 16 de abril de 2016.
Ishaq, S. L., Kim, C. J., Reis, D., and Wright, A. D. G.
(2015). Fibrolytic bacteria isolated from the rumen of North American moose
(Alces alces) and their use as a probiotic in neonatal lambs. Plos One.
10(12): 1-25.
Issakowicz, J., Bueno, M. S., Sampaio, A. C. K., and
Duarte, K. M. R. (2013). Effect of concentrate level and live yeast (Saccharomyces cerevisiae) supplementation on Texel lamb performance and carcass characteristics. Livestock Science. 155(1):
44-52.
Jia, P., Cui, K., Ma, T., Wan, F., Wang, W., Yang, D., ...,
and Diao, Q. (2018). Influence of dietary supplementation with Bacillus
licheniformis and Saccharomycescerevisiae as alternatives to monensin on
growth performance, antioxidant, immunity, ruminal fermentation and microbial
diversity of fattening lambs. Scientific Reports. 8(1): 16712.
Julien, C., Marden, J. P., Auclair, E., Moncoulon, R.,
Cauquil, L., Peyraud, J. L., and Bayourthe, C. (2015). Interaction between live
yeast and dietary rumen degradable protein level: effects on diet utilization
in early-lactating dairy cows. Agricultural Science. 6(1): 1-13.
Kannan, S., Hernandez, L., Herrera, A., Jimenez, B.,
Miller, M., Perales, P., and Subburaj, P. (2014). Genesis of antibiotic
resistance (AR) III: triflingrisk of AR pathogens induced infectious diseases
from regulated concentrated animal feeding operations. The FASEB Journal. 28(1):
986-997.
Kowalik, B., Skomial, J., Miltko, R., and Majewska, M. (2016).
The effect of live Saccharomyces cerevisiae yeast in the diet of rams on
the digestibility of nutrients, nitrogen and mineral retention, and blood serum
biochemical parameters. Turkish Journal of Veterinary & Animal Sciences.
40(5): 534-539.
Krehbiel, C. R., Rust, S. R., Zhang, G., and Gilliland, S.
E. (2003). Bacterial direct-fed microbials in ruminant diets: performance
response and mode of action. Journal of Animal Science. 81(14 supl. 2):
120-132.
Leibtag, E. (2008). Corn prices near record high, but what
about food costs, in Amber Waves. [En línea]. Disponible en:
https://www.ers.usda.gov/amber-waves/2008/february/corn-prices-nearrecord-high-but-what-about-food-costs/.
Fecha de consulta: 16 de septiembre de 2018.
Mahyuddin, P. and Winugroho, M. (2010). Effect of
combination of yeast (Saccharomyces cerevisae + Candida utilis)
and herbs supplementation in finishing
diet on carcass characteristics of beef cattle. Journal
Indonesian Tropical Animal Agriculture. 35(4):
251-256.
Marrero, Y., Castillo, Y., Ruiz, O., Burrola, E., and
Angulo, C. (2014). Feeding of yeast (Candida spp) improves in vitro
ruminal fermentation of fibrous substrates. Journal of Integrative
Agriculture. 14(3): 514-519.
McDonald, I. (1981). A revised model for estimation of protein
degradability in the rumen. Journal of Agricultural
Scienece. 96(1): 251-252.
Mokhber-Dezfouli, M. R., Tajik, P., Bolourchi, M., and
Mahmoudzadeh, H. (2007). Effects of probiotics supplementation in daily milk
intake of newborn calves on body weight gain, body height, diarrhea occurrence
and health condition. Pakistan
Journal of Biological Sciences. 10(18): 3136-3140.
Noziére, P. Y. and Michaelt-Doreau, B. (2000). In sacco
methods. Farm animal metabolism and nutrition (tenth edition.). USA: CABI
Publishing. 438 Pp.
NRC, National Research Council (2001). Nutrientrequeriments
of domestic animals. National Academic Press. Washington, DC: Seventh
edition. 363 Pp.
Obeidat, B. S. (2017). The effects of feeding olive cake
and Saccharomyces cerevisiae supplementation on performance, nutrient
digestibility and blood metabolites of Awassi lambs. Animal Feed Science and
Technology. 231: 131-137.
Obeidat, B. S., Mahmoud, K. Z., Obeidat, M. D., Ata, M.,
Kridli, R. T., Haddad, S. G., ..., and Hatamleh, S. M. (2018). The effects of Saccharomyces
cerevisiae supplementation on intake, nutrient digestibility, and rumen
fluid pH in Awassi female lambs. Veterinary World. 11(7): 1015-1020.
Ørskov, E. R. and Mcdonald, I. (1979). Estimation of
protein degradability in the rumen form incubation measurements weighted
according to rate of passage. Journal of Agricultural Science. 92(2): 499-503.
Promkot, C., Wanapat, M., and Mansathit, J.
(2013). Effects of yeast fermented-cassava chipprotein (YEFECAP) on dietary intake and milk production of Holstein crossbred heifers and cowsduring
pre-and post-partum period. Livestock Science. 154(1-3): 112-116.
Puniya, A. K., Singh, R., and Kamra, D. N. (2015). Rumen
microbiology: From evolution to revolution (primera edición). USA: Springer. 379 Pp.
Qadis, A. Q., Goya, S., Ikuta, K., Yatsu, M., Kimura, A.,
Nakanishi, S., and Sato, S. (2014). Effects of
a bacteria-based probiotic on ruminal pH, volatile fatty acids and bacterial
flora of Holstein calves. Journal of Veterinary Medical Science. 76(6):
877-885.
Ruiz, O., Castillo, Y., Arzola, C., Burrola, E., Salinas, J.,
Corral, A., ..., and Itza, M. (2016). Effects of Candida norvegensis
live cells on in vitro oat straw rumen fermentation. Asian-Australasian
Journal of Animal Sciences.
29(2): 211-218.
Salinas-Chavira, J., Almaguer, L. J., Aguilera-Aceves, C.
E., Zinn, R. A., Mellado, M., and Ruiz-Barrera, O. (2013). Effect of
substitution of sorghum stover with sugarcane top silage on ruminal dry matter
degradability of diets and growth performance of feedlot hair lambs. Small
Ruminat Research. 112(1): 73-77.
SAS, Statistitical Analysis System (2007). User’s Guide:
Statistics, Version 9.6th Edition. SAS Inst., Inc., Cary, NC.
Seo, J. K., Kim, S. W., Kim, M. H., Upadhaya, S. D., Kam,
D. K., and Ha, J. K. (2010). Direct-fed microbials
for ruminant animals. Asian-Australasian Journal of Animal
Sciences. 23(12):
1657-1667.
Shankhpal, S., Parnerkar, S., and Bhanderi, B. M. (2016).
The effect of feeding bypass fat and yeast (saccharomyces cerevisiae)
supplemented total mixed ration on feed intake, digestibility, growth
performance and feed conversion efficiency in weaner Surti kids. Livestock
Research International. 4(1): 11-17.
Tamburello, L., Bulleri, F., Balata, D., and Benedetti-Cecchi,
L. (2014). The role of overgrazing and anthropogenic disturbance in shaping
spatial patterns of distribution of an
invasive seaweed. Journal of Applied Ecology. 51(2): 406-414.
Tristant, D. and Moran, C. A. (2015). The efficacy of
feeding a live probiotic yeast, Yea-Sacc®, on the performance of lactating
dairy cows. Journal of Applied Animal Nutrition. 3(12): 90-95.
Zeoula, L. M., Do-Prado, O. P. P., Geron, L. J. V., Beleze,
J. R. F., Aguiar, S. C., and Maeda, E. M. (2014). Total digestibility and in
situ degradability of bulky diets with the inclusion of ionophores or
probiotics for cattle and buffaloes. Semina: Ciências Agrárias. 35(4):
2063-2076.