https://doi.org/10.29059/cienciauat.v20i1.2017
Fertilización
foliar del frijol con boro y magnesio contra el estrés por altas temperaturas
naturales
Foliar
fertilization of beans with boron and magnesium to prevent high temperature
stress
Fertilización foliar
contra estrés térmico
Alejandro Antonio
Prado-García1,2, Gabriel Iturriaga de-la-Fuente2, César
Leobardo Aguirre-Mancilla2, Jorge Alberto Acosta-Gallegos1*
*Correspondencia:
acosta.jorge@inifap.gob.mx/Fecha de recepción: 10 de abril de 2025/Fecha de
aceptación: 17 de octubre de 2025/Fecha de publicación: 31 de octubre de 2025.
1Centro
Nacional de Investigación disciplinaria (CIR Centro), Campo Experimental Bajío,
Instituto Nacional de Investigaciones Forestales (INIFAP) Agrícolas y
Pecuarias, km. 6.5 carretera Celaya-San Miguel de Allende, Celaya, Guanajuato,
México, C. P. 38110. 2Tecnológico Nacional de México-Instituto
Tecnológico de Roque (TecNM-IT Roque), Celaya, Guanajuato, México, C. P. 38110.
Resumen
El estrés por alta temperatura disminuye el rendimiento y
calidad del frijol. El objetivo del presente trabajo fue determinar el
rendimiento de cuatro variedades de frijol, dos de Phaseolus vulgaris
(Granada y Rarámuri) y dos de Phaseolus acutifolius (TARS-Tep 23 y
Alubia Choix) producidas bajo riego, con la aplicación de boro (B) y magnesio
(Mg) en condiciones de campo con temperaturas naturales altas. Se administraron
en tres ocasiones (una en prefloración y dos durante la floración) cuatro
tratamientos de fertilización foliar: B, Mg, B + Mg y un testigo sin
fertilizar. El ensayo se realizó en marzo de 2020 y 2021. Se cuantificó la
biomasa, peso de 100 semillas, rendimiento e índice de cosecha. Se determinó la
germinación y vigor de la semilla cosechada en cada tratamiento y variedad. En
promedio, P. vulgaris resultó superior (P ≤ 0.05) a P. acutifolius
en peso de 100 semillas y rendimiento. La variedad Rarámuri mostró un mayor
rendimiento, biomasa y peso de 100 semillas.
Los tratamientos de fertilización superaron al testigo, con incremento
promedio de 0.4 g en peso de 100 semillas y 512 kg/ha en rendimiento. La
aplicación foliar de Mg aumentó el rendimiento y el peso de 100 semillas en
ambas especies y variedades evaluadas.
PALABRAS CLAVE: Phaseolus vulgaris L., Phaseolus
acutifolius A. Gray, estrés abiótico, micronutrientes, rendimiento.
ABSTRACT
High temperature
stress lowers bean yield and quality. This work aimed to
determine the yield response of four
bean cultivars, two of P. vulgaris
(Granada and Rarámuri) and two of P.
acutifolius (TARS-Tep 23 and Alubia Choix) grown under irrigation, to the
application of Boron and Magnesium under
field conditions with naturally high temperatures. Four foliar fertilization
treatments were administered on three occasions (one in pre- and two during flowering) B, Mg, B + Mg and a
control group without fertilization. The trial was carried out in 2020 and
2021. Aerial biomass, weight of 100 seeds, yield and the harvest index were
quantified. Germination and vigor of harvested seeds were determined for each
treatment and cultivar. On average, P. vulgaris was superior (P ≤ 0.05)
to P. acutifolius in 100-seed weight and seed yield. The Rarámuri cultivar showed greater yield, biomass, and
100-seed weight. The foliar fertilization treatments surpassed the control
group, with an average increase of 0.4 g in 100-seed weight and 512 kg/ha in
yield. Foliar Mg application increased yield and 100-seed weight in both
species and varieties evaluated.
KEYWORDS: Phaseolus
vulgaris L., Phaseolus acutifolius A. Gray, abiotic stress, micronutrients, yield.

INTRODUCCIÓN
El frijol se cataloga
como un cultivar extremadamente sensible al estrés por calor, principalmente
cuando este ocurre durante la etapa reproductiva inicial (Porch y Jahn, 2001).
El efecto simultáneo de altas temperaturas y déficit de humedad provocan el aborto
de flores y vainas pequeñas, lo que afecta directamente el rendimiento (Porch y Jahn, 2001; Omae y col., 2012). A nivel global, se estima que el cambio
climático incrementará la presencia de sequía y de altas temperaturas, lo que
podría dañar del 60 % al 73 % del área actual de siembra del frijol,
ocasionando pérdidas en la producción del 10 % al 100 % (Beebe y col., 2008).
En El Bajío, el frijol
se produce en dos modalidades de siembra, riego y temporal, siendo más
importantes las primeras, por su alta productividad. Sin embargo, la superficie
bajo riego destinada al cultivo de frijol ha disminuido en los últimos años,
debido a un bajo rendimiento, que podría asociarse a la presencia de
temperaturas superiores a 32 °C, que coinciden con el inicio de la etapa
reproductiva (Anaya-López y col., 2022).
Dichas temperaturas dañan el polen y causan la caída de vainas y un
menor número de semillas en estas (Porch y Jahn, 2001). Anaya-López y col.
(2022) determinaron, a campo abierto, el efecto de la temperatura alta sobre el
rendimiento e índice de cosecha (IC) en germoplasma mejorado de frijol de los
tipos Pinto y Flor de Mayo. En su estudio,
los materiales con bajo IC registraron producción alta de biomasa y un
elevado número de vainas, pero con una o dos semillas, por lo que generaron un
rendimiento inferior. Por el contrario, los
materiales con rendimiento alto mostraron un número elevado de granos
por unidad de área, como resultado de la formación de más vainas con mayor
número de semillas. Omae y col. (2012) obtuvieron resultados similares y
concluyeron que los materiales de frijol tolerantes al calor mostraron alta
capacidad para distribuir fotoasimilados a
las ramas, lo que se tradujo en alta producción de vainas y semillas.
Cakmak
y Yazici (2010) señalaron que, la clorosis en las hojas ocurre bajo condiciones
de estrés abiótico en combinación con una deficiencia nutrimental de potasio
(K) y Mg. Indicaron que las plantas con deficiencia de Mg fueron más
susceptibles al estrés por altas temperaturas, aumentando la clorosis, ya que
este elemento constituye 2.7 % de la clorofila y es fundamental en la
fotosíntesis, además, promueve el crecimiento radicular y el transporte de
fotoasimilados en la planta. Adicionalmente, el Mg actúa como cofactor en más
de 300 reacciones enzimáticas críticas bajo estrés térmico, incluyendo la
síntesis de adenosín trifosfato (ATP) y la activación de la
ribulosa-1,5-bifosfato carboxilasa/oxigenasa (RuBisCO), enzimas todas, que se
ven severamente comprometidas por las altas temperaturas (Baloch y col., 2022).
El boro (B) es otro elemento involucrado en múltiples funciones como la
síntesis y estructura de la pared celular, lignificación, metabolismo y
transporte de carbohidratos e integridad de la membrana plasmática, entre otros
procesos que se ven afectados por las altas temperaturas (Kirkby y Rómheld,
2008). Se estima que cerca del 90 % del B de la planta se encuentra en las
paredes celulares, por lo cual puede considerarse como un elemento fundamental
para contrarrestar los efectos provocados por las altas temperaturas (Ahmad y
col., 2009). La deficiencia de dicho elemento afecta la formación del tubo
polínico y con ello la formación del fruto, lo que conlleva a bajos
rendimientos. Además, el B es fundamental para la germinación del polen y el
crecimiento del tubo polínico, procesos extremadamente sensibles a las altas
temperaturas y que pueden determinar el éxito reproductivo del cultivo bajo
estrés térmico (Behera y col., 2023). Este problema se puede abordar con el uso
de variedades tolerantes a altas temperaturas, en combinación con la nutrición
vegetal adecuada (Anaya-López y col., 2022). Por otra parte, entre las especies
domesticadas del género Phaseolus, el frijol Tepari o escomite es
mencionado en la literatura como tolerante a altas temperaturas (Rainey y
Griffiths, 2005).
El objetivo de este
estudio fue determinar el rendimiento y la calidad fisiológica de la semilla de
cuatro variedades de frijol, dos de P. vulgaris y dos de P.
acutifolius, en respuesta a la aplicación foliar de B y Mg, en condiciones
de altas temperaturas naturales.
MATERIALES Y MÉTODOS
Sitio de estudio
El
estudio se realizó bajo condiciones de campo abierto en el Campo Experimental Bajío (CEBAJ) del (Instituto Nacional de Investigaciones
Forestales, Agrícolas y Pecuarias) (INIFAP), localizado a los 20º34'
47'' N, 100º49'22'' W y 1 764 msnm. El clima predominante de la zona es BS1hw
(W). La temperatura promedio oscila en los 20.6 °C y la precipitación anual promedio es de 597 mm (Mora y col., 2001).
Material vegetal
Se utilizaron cuatro
variedades de frijol de dos especies: P. vulgaris L. o frijol común,
variedades locales de Granada y Rarámuri, y P. acutifolius o frijol
Tepari, variedades TARS-Tep 23 (Puerto Rico) y Alubia Choix, Sinaloa.
Diseño experimental
Estudio agronómico
Para las evaluaciones
de campo se utilizó un diseño de bloques completos al azar con arreglo de
parcelas divididas y tres repeticiones en cada ciclo de cultivo. Este diseño se
empleó para controlar el error experimental inherente a las condiciones heterogéneas
de campo. La parcela grande estuvo constituida por las cuatro variedades y la
subparcela por los cuatro tratamientos de fertilización foliar. Los
tratamientos de fertilización fueron YaraVita Boro® al 11 %, 0.5 L/ha (por
unidad experimental se aplicaron 0.57 mL/1 L de agua); YaraVita Magnesio®
al 20 %, 4.0 L/ha (por unidad experimental se administraron 4.56 mL/1 L de
agua); B + Mg (por unidad experimental se aplicaron 0.57 mL de B + 4.56
mL de Mg/1 L de agua) y un testigo sin fertilizante.
Aplicación
de tratamientos foliares
Los tratamientos de
fertilización foliar se administraron en tres ocasiones durante el ciclo de
cultivo: la primera se realizó en la etapa de prefloración (aproximadamente a
los 35 DDS después de la siembra, DDS), y las
dos aplicaciones restantes durante la floración (a los 45 DDS y 55 DDS,
aproximadamente). Los tratamientos se llevaron a cabo mediante aspersión
foliar, con bomba de mochila (Urrea, FUMU20, El Salto, Jalisco, México),
aplicando un volumen de 300 L/ha en las primeras horas de la mañana (7: 00 h a
9: 00 h), para minimizar la evaporación y maximizar la absorción foliar. Se
utilizó agua como vehículo y se añadió
surfactante no iónico (0.1 % v/v) para mejorar la adherencia y
penetración de los micronutrientes en las hojas.
Manejo agronómico
La siembra se realizó
a tierra venida, después de un riego de preparación, con una lámina de 30 mm.
Las fechas de siembra fueron el 16 de marzo de 2020 y 13 de marzo de 2021. El
manejo agronómico fue el mismo en ambos
ciclos. El suelo se fertilizó con la dosis 80-60-40 de N-P-K. Se
establecieron 12 plantas/m/L en surcos a 76 cm de distancia, lo que representa
una densidad de población de aproximadamente
157 895 plantas/ha. La parcela de cada unidad experimental estuvo
constituida por 3 surcos de 5 m de longitud. Durante la etapa vegetativa se
dieron 2 cultivos mecánicos para controlar maleza. Se llevó a cabo un segundo
riego rodado con una lámina de 30 mm a los 30
d después de la siembra (DDS). A los 65 DDS se colocó cintilla para riego por goteo con un gasto de 1 L/h. A
partir de ese momento se realizaron 3 riegos semanales, aplicando en cada
ocasión una lámina de 15 mm desde los 65 DDS hasta los 95 DDS (195 mm). La lámina total de riego suministrada durante el
ciclo de cultivo fue de aproximadamente 255 mm.
Variables evaluadas
Las variables se
evaluaron en parcelas útiles (a las cuales se eliminó el efecto de borde para
asegurar que todas las plantas tuvieron competencia uniforme con sus vecinas)
de un surco central de 5 m/L. Los días a floración se registraron cuando el 50
% de las plantas presentó al menos una flor abierta. La altura de planta se
midió desde la base del tallo hasta el ápice en 5 plantas tomadas al azar
dentro de la parcela útil. El número de vainas por planta y el número de
semillas por vaina se determinaron a partir de 5 plantas individuales tomadas
al azar dentro de la parcela útil.
El
rendimiento de semilla (kg/ha) se obtuvo cosechando la parcela útil completa y
ajustando la humedad al 14 % mediante la fórmula:
Rendimiento
ajustado = [Peso húmedo × (100 - humedad inicial)] ÷ (100 - 14)
El peso de 100
semillas (g) se determinó contando y pesando 100 semillas tomadas al azar de
cada parcela.
Índice de cosecha (IC)
Se obtuvo un IC
modificado, mediante la recolección del material biológico presente en 0.5 m/L
antes de la senescencia de las hojas, procurando recolectar en bolsas todas las
hojas caídas previo al muestreo. En el cálculo de biomasa aérea se incluyó el peso
de tallos, vainas (sin semillas) y hojas. En el caso de las semillas se incluyó
solo el peso de las semillas presentes en la muestra recolectada, sin las
vainas. El IC se calculó utilizando la siguiente fórmula:
IC =
[(Peso de semilla/Peso de biomasa aérea total) × 100]
Durante los dos ciclos
del cultivo de frijol de riego, se registró la temperatura y precipitación
pluvial cada tercer día, en una estación meteorológica automática (Vantage
Pro2, Davis Instruments, Hayward, CA, EE. UU.), ubicada a 300 m del sitio
experimental.
Caracterización de las
semillas
Las semillas obtenidas
de cada tratamiento de campo fueron evaluadas en el laboratorio para determinar
las variables de calidad fisiológica del porcentaje de germinación estándar a
los 8 d, emergencia de radícula a los 4 d (vigor), peso fresco y seco de plántula,
y número de semillas muertas y anormales. Las pruebas se realizaron bajo
condiciones controladas de temperatura (25 °C ± 2 °C) y humedad relativa (65 %
± 5 %) utilizando 50 semillas por tratamiento con cuatro repeticiones.
Análisis estadístico
El análisis
estadístico de cada ciclo individual se realizó mediante un diseño de bloques
completos al azar con arreglo de parcelas divididas, donde la parcela principal
correspondió a las variedades y la subparcela a los tratamientos de
fertilización foliar, incluyendo sus interacciones.
En el análisis combinado de ambos ciclos (2020 y 2021) se empleó un
diseño factorial generalizado con los
factores año, variedad, fertilización foliar y sus interacciones.
Adicionalmente,
se llevaron a cabo análisis multivariados
(Componentes Principales y Cluster Jerárquico
Aglomerativo) utilizando los promedios de los tres factores principales. Todos
los análisis estadísticos se realizaron mediante el paquete estadístico SAS
v9.4 (SAS Institute Inc., Cary, NC, EUA) y Genotype-Environment Analysis in R (GEA-R) v4.1 (Desarrollador: Centro
Internacional de Mejoramiento de Maíz y Trigo (CIMMYT) (Texcoco, México); los
análisis multivariados complementarios,
gráficos y cálculos se elaboraron en Microsoft Excel 2016 versión 16.0
(Microsoft Office, Redmond, WA, EUA).
Análisis de
correlación
Las variables
agronómicas se evaluaron independientemente
del tratamiento de fertilización aplicado.
Para la interpretación de las correlaciones se utilizó la escala de
Cohen (1988): débil (r = 0.10 a 0.29), moderada (r = 0.30 a 0.49), fuerte (r =
0.50 a 0.70) y muy fuerte (r > 0.70).
Análisis de semilla
por efecto de genotipo y tratamiento de fertilización foliar
Para las evaluaciones
de germinación y vigor de la semilla, se utilizó un diseño completamente al
azar con arreglo factorial, ya que las condiciones controladas de laboratorio
no requieren bloqueo para el control del error experimental. Los factores del arreglo
factorial fueron las cuatro variedades (genotipos) y los cuatro tratamientos de
fertilización foliar aplicados por triplicado durante la producción de la
semilla en campo.
Se utilizaron semillas
provenientes de plantas del ensayo de campo donde se evaluaron los cuatro
genotipos bajo diferentes tratamientos de fertilización foliar (B, Mg, B + Mg y
testigo), para analizarlas por efecto de genotipo y tratamiento de fertilización
foliar.
RESULTADOS Y DISCUSIÓN
Temperaturas máximas y
mínimas en el ciclo
Las temperaturas
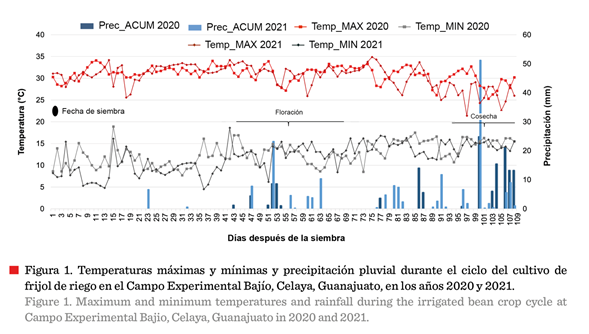
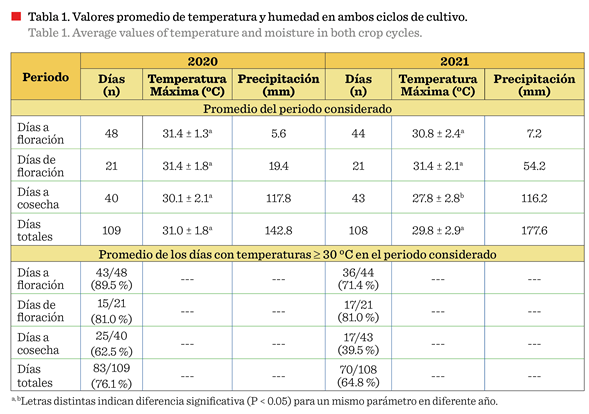
máximas reportadas en la zona de cultivo (Figura 1; Tabla 1), durante la etapa
crítica de floración (21 d) de ambos ciclos (2020 y 2021), oscilaron entre 21.4
°C y 34.9 °C, mientras que las mínimas
variaron entre » 4.5 °C y » 18.9 °C. Se
registraron 17 d de temperaturas máximas consistentemente ≥ 30 °C en los
2 años. Aunque no se calculó la duración en horas de estas temperaturas
elevadas, estuvieron constantemente presentes, toda vez que
en 2020, se registraron hasta 7 d consecutivos de temperatura ≥ 30 °C; mientras
que en 2021, se reportaron
hasta 5 d consecutivos con estas mismas
condiciones climáticas, la cuales fueron críticas para los cultivos. La
exposición a temperaturas entre 28 °C y 32 °C durante
la etapa reproductiva disminuye el número de vainas y semillas,
afectando el tamaño de estas últimas, ya que la temperatura es uno de los parámetros ambientales con mayor influencia en el desarrollo del frijol durante todo
su ciclo de cultivo (Barrios-Gómez y López-Castañeda, 2009).
Efecto del año
Durante
el periodo de siembra 2020, el 76.1 % de los 108 DDS presentaron temperaturas ≥ 30 ºC, en tanto que, durante el periodo 2021, el
64.8 % de los DDS registraron ese comportamiento
(Figura 1; Tabla 1).
En 2020, la etapa
vegetativa tuvo mayor duración (48 d a floración) que en 2021 (44 d a
floración) (Tabla 1). Esto pudo haber sido influenciado por las temperaturas
mínimas que se presentaron en 2020 (7.7 °C a 18.9 °C) respecto a las de 2021
(4.5 °C a 18.6 °C), aunado a la menor precipitación (5.6 mm vs 15.0 mm) así como a la ausencia de precipitaciones
significativas durante el establecimiento del cultivo (Figura 1).
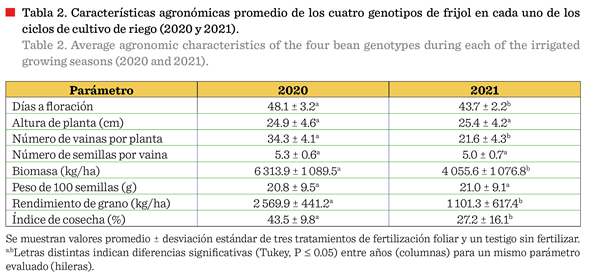
La altura de planta no
varió por efecto del ciclo de siembra, pero la biomasa fue significativamente
mayor (P £ 0.05) en 2020 (Tabla 2). Dicha acumulación de biomasa podría deberse a que, las condiciones climáticas iniciales
de 2020 (89.5 % de temperaturas ≥ 30) (Tabla
1) pudieron haber permitido un desarrollo
radicular más profundo y robusto, usualmente asociado con mayor
producción vegetativa (Tapia y col., 2022) y explicaría el mayor número de
vainas por planta (P £ 0.05) (Tabla 2). Esta característica en particular
estuvo asociada al rendimiento superior obtenido en 2020, y con ello, el mayor
IC (ambos con P £ 0.05 respecto al 2021), ya que el número de semillas por
vaina y el peso de 100 semillas no mostraron diferencia.
Efecto por variedad
El
peso de 100 semillas reportó diferencia (P £ 0.05) entre los
cuatro tratamientos. En P. acutifolius varió de 11.6 g a 13.0 g, en
tanto que P. vulgaris registró entre 26.1 g y 32.8 g (Tabla 3). Se ha
reportado que las plantas y semillas de P. acutifolius son
intrínsecamente de menor tamaño y peso en comparación con las de P. vulgaris, pero también que estas
variedades presentan mayor tolerancia a
estrés hídrico y térmico (Rainey y Griffiths, 2005). Si bien, la comparación entre especies diferentes tiene limitaciones,
es importante su análisis para evaluar alternativas de cultivo, bajo las condiciones locales del
estudio. Asociado a esta diferencia
genotípica, el potencial de rendimiento entre ambas especies fue diferente (P £ 0.05), resultando P. vulgaris
superior a P. acutifolius (Tabla 3). Las variedades de P.
vulgaris (Granada y Rarámuri) registraron rendimientos entre 1 972.8 kg/ha y 2 207.5 kg/ha, mientras que los Teparies (TARS-Tep
23 y Alubia Choix) reportaron rendimientos entre
1 504.7 kg/ha y 1 657.7 kg/ha, representando aproximadamente 65 % a 75 % del rendimiento
del frijol común. Estas diferencias son congruentes con reportes previos
que indican menor potencial productivo pero
mayor tolerancia al estrés en P. acutifolius (Rainey y Griffiths,
2005), el cual mantuvo un rendimiento relativamente estable
(diferencia de solo 153 kg/ha entre variedades) comparado con la mayor
variabilidad en P. vulgaris (diferencia de 235 kg/ha entre variedades),
lo que podría indicar mayor consistencia bajo estrés térmico (Tabla 3).
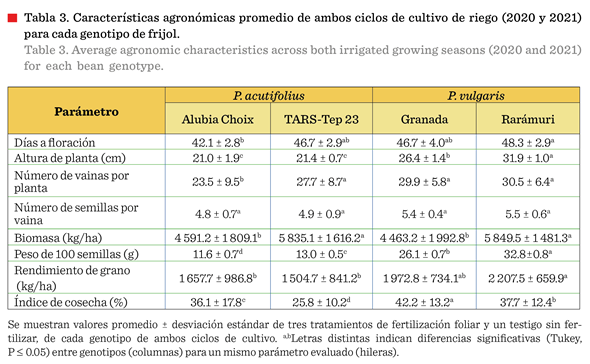
Entre los Teparies,
Alubia Choix presentó mayor IC (36.1 % vs 25.8 %) (P £ 0.05) que TARS-Tep 23
(Tabla 3), característica probablemente asociada a una mejor resistencia al
estrés térmico tardío. Entre las variedades de P. vulgaris, Rarámuri
mostró superioridad (P £ 0.05) en altura de planta (31.9 cm vs 26.4 cm),
biomasa (5 849 kg/ha vs 4 463 kg/ha) y peso de 100 semillas (32.8 g vs 26.1 g),
respecto a la variedad Granada, pero un menor IC
(37.7 % vs 42.2 %) (P £ 0.05) (Tabla 3). Ello se explica por la diferencia
en el peso de las semillas entre ambas variedades (un
20.4 % más de peso por semilla), lo
que implica que Rarámuri requiere
mayor inversión de fotoasimilados por semilla individual, resultando en un menor IC a pesar de su superior
rendimiento total. Adicionalmente, Rarámuri presentó el mayor número de vainas por planta (30.5) y semillas por vaina (5.5), características directamente
relacionadas con su superior rendimiento, aunado a que dicha variedad de
material de tipo pinto con alto potencial de rendimiento y respuesta neutral al
fotoperiodo (Anaya-López col., 2022), está bien adaptado a la localidad de
prueba, donde se desarrolló.
Los resultados
obtenidos señalan las diferencias en las características fisiológicas y
morfológicas de las plantas que son conducentes a la obtención de altos
rendimientos entre el frijol Tepari y el frijol común bajo condiciones de
estrés ambiental.
Tratamiento de
nutrición foliar (B y Mg)
Los tratamientos con
fertilización foliar, aplicando micronutrientes superaron al testigo, con un
incremento promedio de 0.4 g en peso de 100 semillas y de 512 kg/ha en
rendimiento (Tabla 4).
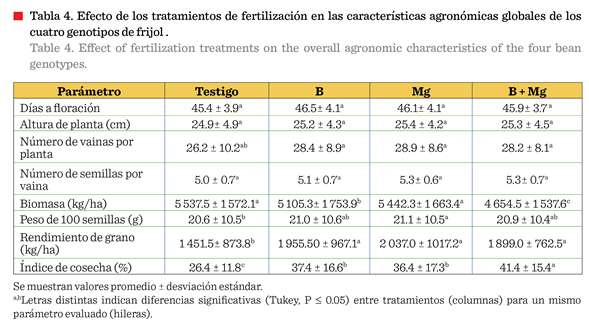
El incremento de
rendimiento varió de 40.3 %, 34.7 % y 30.8 % para Mg, B y B + Mg,
respectivamente. La aplicación de fertilización foliar mejoró consistentemente
el IC en todos los genotipos, siendo este el parámetro más sensible a los
tratamientos de micronutrientes. El Mg elevó el rendimiento y el peso de 100
semillas en ambas especies y variedades evaluadas con fertilización foliar.
Los
tratamientos individuales de B y Mg, mostraron
efectos similares a la combinación B + Mg en la mayoría de los diferentes
aspectos agronómicos, aunque la mezcla presentó mayores valores (P ≤ 0.05) de
biomasa e IC (Tabla 4). Sin embargo, este efecto de mejora fue distinto para
las dos especies (Tabla 5), sugiriendo que no existe un efecto sinérgico entre estos micronutrientes bajo las condiciones
del estudio. El que la aplicación de los dos
elementos juntos, B + Mg, no mostrase una ventaja consistente sobre su
administración individual en las diferentes características evaluadas, sugiere
posibles interacciones antagónicas, o que
las dosis óptimas de cada elemento cambian cuando se aplican en
combinación.
Siddiqui
y col. (2018) señalaron que el Mg ayuda
en la protección de las plantas contra el estrés por calor al aumentar el área
foliar, lo que resulta en mayor materia seca. Stagnari y col. (2009) estudiaron
el efecto de la fertilización foliar de Mg (56 g/ha, 112 g/ha y 224 g/ha en
aplicación única en la floración o dividiendo la administración en dos etapas,
la mitad en etapa de cuatro hojas y la mitad en la floración) en dos genotipos
de frijol ejotero, Bronco y Cadillac, y observaron que la fertilización en una
sola dosis, incrementó de manera significativa el rendimiento de vainas
frescas, en cada una de las 2 variedades
estudiadas (78 % y 32 %, respectivamente). La aplicación dividida de Mg
en dos etapas también fue efectiva al elevar el rendimiento en 109 % y 50 %
para los 2 genotipos estudiados, respectivamente.
El incremento en las
variables agronómicas por la adición de los minerales solos o combinados es
relevante, considerando que, bajo estrés térmico severo, mantener o mejorar
ligeramente el peso de semillas representa una ventaja adaptativa importante,
ya que normalmente el estrés por calor reduce significativamente este parámetro
(Porch y Jahn, 2001). En términos prácticos,
dicho aumento podría traducirse en mejor calidad comercial y vigor de la semilla, aspectos críticos para la
productividad en ciclos subsecuentes bajo condiciones adversas, lo
anterior podría deberse a la protección que aporta ese elemento al polen,
principalmente en la etapa reproductiva, etapa extremadamente sensible al
estrés por calor (Porch y Jahn, 2001). Un
balance adecuado en la nutrición de los cultivos influye positivamente
en su desarrollo vegetativo y re-productivo (Feng y col., 2023). Los resultados
obtenidos muestran respuesta positiva en los parámetros
de rendimiento para ambos nutrientes
en los genotipos de frijol evaluados bajo las condiciones de estrés
térmico del estudio.
Un hallazgo destacado
fue que el testigo reportó la mayor biomasa (5 537.5 kg/ha) pero el menor IC (26.4 %), mientras que los tratamientos
con fertilización foliar (B y/o Mg), presentaron menor biomasa, pero
mayor eficiencia en la partición de asimilados hacia el grano (IC: 37.8 % a
41.4 %) (Tabla 4). Esto sugiere que, bajo
estrés térmico, las plantas sin fertilización foliar destinan mayor proporción
de fotoasimilados al crecimiento vegetativo como
mecanismo de supervivencia, reduciendo la inversión reproductiva. La
aplicación de B y Mg mejoró la eficiencia reproductiva probablemente al
optimizar procesos críticos como la
formación del tubo polínico (B) y la actividad enzimática fotosintética
(Mg), favoreciendo la translocación
de carbohidratos hacia los órganos
reproductivos. Este fenómeno es consistente
con lo reportado por Polania y col.
(2016) en condiciones de estrés abiótico, donde la nutrición balanceada
mejoró la partición de asimilados hacia el
rendimiento económico.
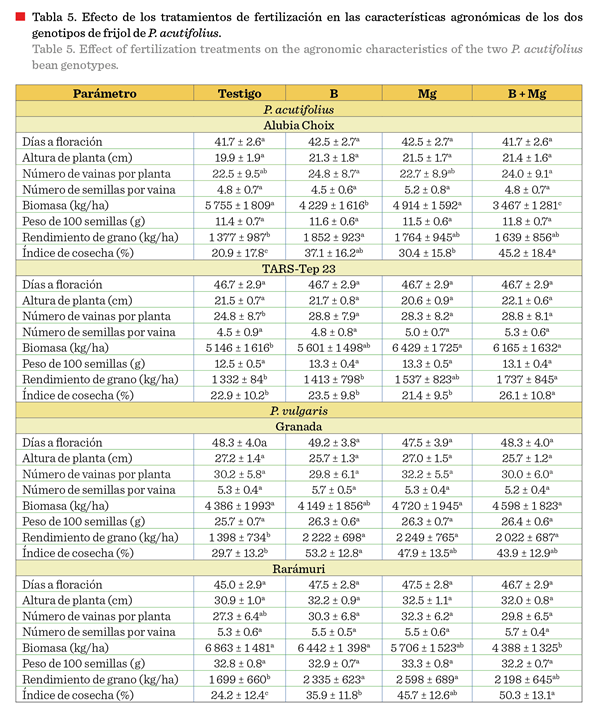
Los tratamientos con fertilización foliar en cada
genotipo revelaron respuestas diferenciales entre especies y variedades
(Tabla 5). En la especie de P. vulgaris, el testigo de Granada mostró rendimientos de 1 398 kg/ha
que fueron superados por los 3 tratamientos, siendo el de Mg el que obtuvo el
mayor valor con 2 249 kg/ha; en tanto que,
en Rarámuri, el testigo presentó 1 699 kg/ha, valor que fue superado por los tratamientos de B y Mg, siendo este
último el de mayor rendimiento (2 598 kg/ha). En
contraste, los genotipos de P. acutifolius mostraron menor respuesta, sin embargo, tuvieron
el mismo comportamiento que P. vulgaris; el testigo de Alubia Choix registró el valor inferior con 1 377 kg/ha; B
reportó el valor máximo con 1 852 kg/ha. En
TARS-Tep 23, el testigo obtuvo 1 332 kg/ha y B + Mg el mayor valor (1
737 kg/ha). Estos resultados evidencian que P. vulgaris fue más
responsivo a la aplicación de
micronutrientes que P. acutifolius, y ambos genotipos
presentaron respuesta diferenciada.
La mejora en el
rendimiento se asoció con características como mayor número de vainas por
planta (Rarámuri: 32.3, 30.3 y 29.8 vainas para Mg, B y B + Mg vs 27.3 vainas
en el testigo) y mayor IC (45.7 %, 35.9 % y 50.3 % ,
respectivamente, vs 24.2 % en el testigo). Altos valores de IC se asociaron con
la obtención de rendimientos superiores en los tratamientos con fertilización
foliar, lo que sugiere un efecto de los nutrientes aplicados en la asignación de asimilados a las estructuras reproductivas, aunque paradójicamente el
testigo produjo la mayor biomasa total (6 863 kg/ha). Ello indica que,
los micronutrientes mejoraron la eficiencia de partición hacia grano más que la
producción total de biomasa. Siddiqui y col.
(2018), reportaron resultados en Vicia faba, donde encontraron
que la aplicación de Mg a la planta bajo condiciones de estrés por calor
aumenta significativamente hasta un 53.14 % el área foliar, además de elevar la
concentración de osmoprotectores como prolina (PRO) y glicina betaína (GB) en
un 43.31 % y 47.10 %, respectivamente, en comparación con el testigo sin estrés
por altas temperaturas.
En P. acutifolius,
los tratamientos con fertilización foliar mejoraron significativamente el
rendimiento de grano comparado con el testigo, especialmente en Alubia Choix,
donde el tratamiento con B incrementó el rendimiento de 1 377 kg/ha a 1 852 kg/ha. El IC mostró las mejores respuestas,
alcanzando valores superiores con la combinación B + Mg (45.2 %) en Alubia
Choix, comparado con el testigo (20.9 %). En TARS-Tep 23, la aplicación
de Mg favoreció significativamente la acumulación de biomasa (6 429 kg/ha vs 5
146 kg/ha del testigo), aunque el rendimiento de grano se mantuvo
relativamente bajo en comparación con P. vulgaris. En P.
vulgaris, la respuesta a la fertilización foliar fue más pronunciada.
Granada mostró incrementos notables en rendimiento con los tratamientos B y Mg (2 222 kg/ha y 2 249 kg/ha,
respectivamente) comparado con el testigo (1
398 kg/ha). El IC en Granada aumentó significativamente con la aplicación de B
(53.2 % vs 29.7 % del testigo), indicando una mejor
partición de fotoasimilados hacia estructuras reproductivas. Rarámuri, la variedad de mayor rendimiento,
respondió favorablemente a todos los tratamientos, alcanzando su mejor desempeño con Mg (2 598 kg/ha) y manteniendo
el mayor IC con B + Mg (50.3 %).
Ambas
variedades de frijol mostraron mejoras sustanciales en el IC con los tratamientos de
micronutrientes, confirmando la eficiencia
de la fertilización foliar en la partición de asimilados hacia órganos reproductivos bajo condiciones de estrés
térmico.
Análisis de
correlación
El análisis de
correlación general reveló las interrelaciones
entre las variables agronómicas evaluadas, independientemente del tratamiento de fertilización aplicado (Tabla 6). Los
días a la floración mostraron correlaciones
muy fuertes con la altura de planta (r
= 0.93***), el peso de 100 semillas (r = 0.94***) y número de vainas por planta
(0.82***); fuertes con el rendimiento (r = 0.52***) y la biomasa (r =
0.55***); y correlación débil con el IC (r = 0.21*). Esto indica que los genotipos con floración más tardía
tendieron a tener plantas con mayor altura, producir semillas de mayor peso y mayor rendimiento, posiblemente por evadir parcialmente el
estrés térmico temprano o por tener mayor tasa de llenado de vainas.
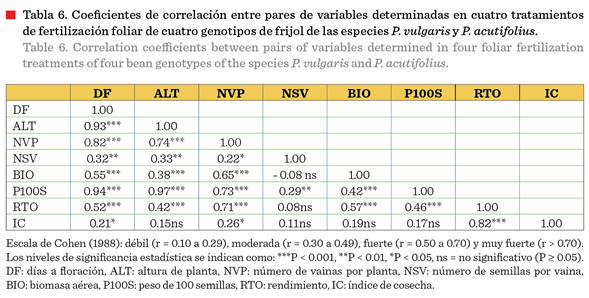
La
altura de planta mostró correlaciones muy fuertes con el peso de 100 semillas (r = 0.97***) y con el número de vainas por planta (r =
0.74***), confirmando que plantas más altas tendieron a acumular mayor
materia seca y producir más vainas. También reportó correlaciones moderadas con
el rendimiento (r = 0.42***), biomasa (r = 0.38***) y número de semillas por
vaina (r = 0.33**). El número de vainas por planta presentó
correlaciones muy fuertes con el peso de 100
semillas (r = 0.73***) y rendimiento (r = 0.71***), mientras que con
biomasa tuvo una correlación moderada
(0.65***). Esto muestra al número de vainas como un componente
importante en la productividad del frijol, tal y como lo observaron Anaya-López y col. (2022) en la misma
localidad de prueba.
La biomasa presentó
correlaciones positivas fuertes con el
rendimiento (r = 0.57***), y moderadas con el peso de 100 semillas (r =
0.42***), indicando que la capacidad de acumulación de materia seca
contribuye al rendimiento final.
El rendimiento mostró
una correlación positiva muy fuerte con el IC (r = 0.82***), confirmando su
papel como indicador de eficiencia en la
partición de fotoasimilados hacia estructuras reproductivas.
Sin embargo, el IC
tuvo correlaciones no significativas con la biomasa (r = 0.19), el peso de 100
semillas (r = 0.17), el número de semillas por vaina (r = 0.11) y con altura de
planta (r = 0.15).
Estas correlaciones
generales proporcionan el contexto para interpretar cómo los tratamientos de fertilización foliar influyeron en las
variables de respuesta, considerando sus interrelaciones naturales en el
germoplasma evaluado.
Interacción genotipo
por tratamiento
El análisis de la
interacción entre genotipos y tratamientos de fertilización foliar, para el
rendimiento, mostró que los dos primeros componentes principales explicaron el
98.62 % de la variación total (CP1 = 94.14 % y CP2 = 4.48 %), siendo el primer
componente el que explicó la mayor proporción (Figura 2). El CP1 se asoció
principalmente con el rendimiento del grano, mientras que el CP2 reflejó la
respuesta diferencial a los tratamientos de fertilización foliar. En el biplot
se observa la respuesta diferencial de los genotipos según su especie: los
genotipos de P. vulgaris (Rarámuri y Granada) se ubicaron en el sector
positivo del CP1 y mostraron respuesta positiva a la aplicación de
fertilización foliar con B y Mg, mientras que los genotipos de P.
acutifolius (Alubia Choix y TARS-Tep 23) presentaron una respuesta
diferente a estos tratamientos.
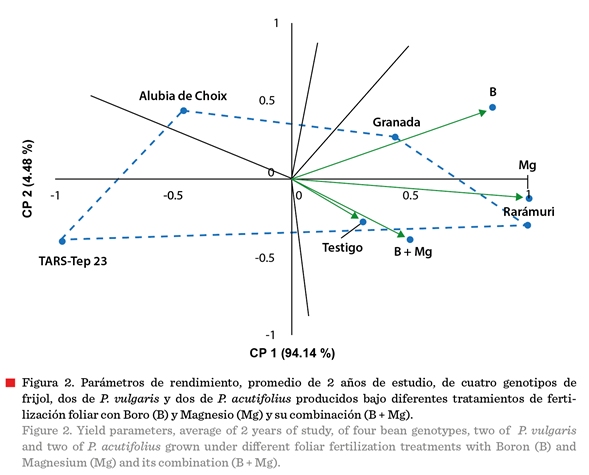
En el cuadrante
inferior derecho (valores positivos de CP1 y CP2) se ubicaron Rarámuri junto
con los tratamientos Mg y B + Mg, indicando la mayor afinidad de esta variedad
de P. vulgaris con la fertilización foliar. Granada se posicionó en el
cuadrante superior derecho, mostrando también
respuesta favorable pero diferente a Rarámuri (Mg y B). Los tratamientos
B y Mg se posicionaron muy próximos entre sí en el biplot, sugiriendo efectos
similares sobre el rendimiento.
El tratamiento testigo
se localizó en el cuadrante inferior derecho, aunque más cercano a la parte de
la recta numérica negativa vertical, y al 0 de la horizontal, lo que indica
diferencias en los requerimientos nutrimentales, ya que está próximo a B + Mg.
Los genotipos de P. acutifolius
(Alubia Choix y TARS-Tep 23) también se distribuyeron en diferentes
cuadrantes, indicando respuestas variables entre ellos. TARS-Tep 23 se ubicó en el cuadrante inferior izquierdo, lo que sugiere
diferencias en los requerimientos
nutricionales y/o mecanismos de absorción y utilización de micronutrientes entre especies. La fertilización combinada
B + Mg se posicionó cerca del centro del biplot, indicando que mostró un efecto
intermedio comparado con las aplicaciones individuales.
La respuesta
diferencial de P. acutifolius a la fertilización foliar puede explicarse
por diferencias intrínsecas en los mecanismos de absorción y translocación de
micronutrientes entre especies. El frijol Tepari, adaptado evolutivamente a
condiciones áridas con suelos de baja fertilidad, podría presentar menor capacidad de absorción foliar de B y Mg, o bien, sistemas
de transporte internos menos eficientes para estos elementos (Rainey y
Griffiths, 2005). Alternativamente, es posible que P. acutifolius requiera
concentraciones o formulaciones diferentes de estos micronutrientes, o que su
metabolismo celular bajo estrés térmico no
se beneficie de la misma manera que P. vulgaris, de la aplicación
exógena de estos elementos (Bornowski y col., 2023).
La posición de
Rarámuri, en el extremo positivo del CP1 refleja su superioridad en rendimiento
y respuesta a fertilización foliar (Figura 2). Estos resultados de la variedad
Rarámuri se asociaron con el mayor rendimiento de grano, altura de planta,
biomasa y peso de 100 semillas (Tabla 5), variables estrechamente ligadas con
el rendimiento, como se confirmó en el análisis de correlación (Tabla 6). Klaedtke y col. (2012) reportaron que
una mayor altura de planta está involucrada en una producción superior de biomasa, y este
volumen de biomasa, puede significar mayor disponibilidad de fotoasimilados
para la removilización y asignación de reservas de los tallos y ramas hacia las
vainas y semillas, mejorando así los rendimientos bajo condiciones de estrés
abiótico. Tene y col. (2023) analizaron la
resistencia a calor de una amplia muestra
de germoplasma de P. vulgaris y de otras especies del género Phaseolus,
y señalaron que, aunque la mayoría de los genotipos estudiados de las especies P.
acutifolius, P. lunatus y P. coccineus, resultaron más
resistentes al calor que la mayoría de los genotipos de P. vulgaris,
algunos genotipos de frijol común mostraron una respuesta similar a la de esas especies. Los resultados del presente
trabajo confirman esta variabilidad
intraespecífica, ya que Rarámuri tuvo una mejor adaptación relativa al estrés térmico comparado con Granada,
evidenciado en su superior rendimiento bajo las condiciones del estudio.
En su investigación, Tene y col. (2023), entre otras características agronómicas,
encontraron que la biomasa mostró los efectos directos más altos en las
calificaciones de resistencia al estrés por calor, lo que es consistente con
los resultados obtenidos en este estudio, donde la mayor biomasa de Rarámuri se
asoció con su superior rendimiento.
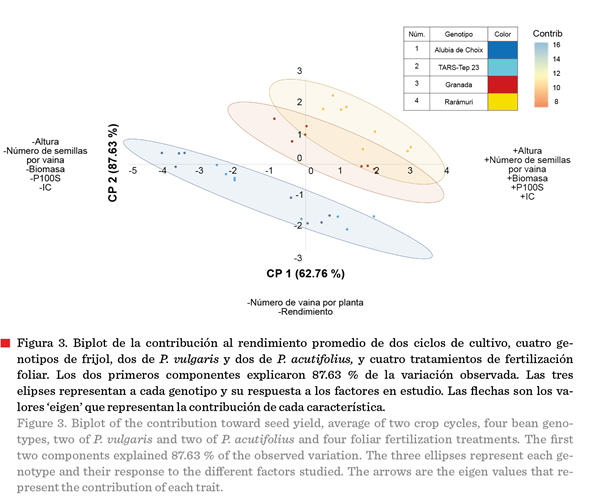
Análisis de
componentes principales
El análisis de
componentes principales de las variables agronómicas (Tabla 7) mostró que los primeros dos componentes explicaron el 87.63 % de la variación total (Figura 3). El CP1 con el 62.76 % de la variación estuvo definido
principalmente por las variables número de vainas por planta y rendimiento, variables que entre sí presentaron
una correlación positiva moderada (r = 0.71***) (Tabla 6).
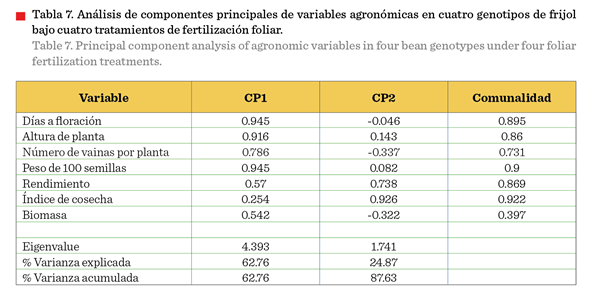
El CP2, con una
aportación a la varianza total del 24.87 %, se constituyó principalmente por las variables altura, número de semillas por
vaina, biomasa, peso de 100 semillas (Figura 3), rasgos que también mostraron asociación positiva en el
análisis de correlación (Tabla 6).
Esta separación en dos
componentes principales indica que el rendimiento está determinado por dos
estrategias principales: una basada en componentes reproductivos (vainas,
semillas, días a floración) y otra en vigor vegetativo (altura, biomasa, peso
de semillas).
Los resultados del
análisis biplot (Figura 3) y los valores del CP1 con mayor contribución a la
variabilidad total permitieron separar la población de datos en tres grupos
distintos, como se observa en las elipses de confianza. El grupo superior
(elipse amarilla) corresponde principalmente a los genotipos de P. vulgaris (Rarámuri
y Granada) bajo tratamientos de fertilización foliar, caracterizados por
valores positivos en ambos componentes principales. El grupo central (elipse
naranja) incluye a los genotipos de P. vulgaris bajo condiciones de
testigo, con valores intermedios. El grupo
inferior (elipse azul) agrupa a los genotipos de P. acutifolius (Alubia
Choix y TARS-Tep 23) bajo todos los tratamientos, ubicados en valores
negativos del CP1. A pesar de provenir de sitios geográficos diferentes, uno de
México y otro de Puerto Rico, estas variedades mostraron comportamiento similar
bajo las condiciones del estudio. La clara separación en tres grupos refleja
las diferencias intrínsecas entre especies y la respuesta diferencial a la
fertilización foliar. En la localidad de prueba, representativa de la región de
El Bajío, las variedades de P. vulgaris resultaron superiores a las de
P. acutifolius en la mayoría de las variables evaluadas, especialmente
cuando se aplicaron tratamientos de fertilización
foliar, confirmando la mejor adaptación del frijol común a estas condiciones
agroecológicas y su mayor respuesta a la nutrición
foliar. La respuesta superior de los genotipos de P. vulgaris
sobre los de P. acutifolius en condiciones de altas temperaturas,
contradice la percepción tradicional que señala al frijol Tepari como más
tolerante al calor (Rainey y Griffiths, 2005). Este hallazgo sugiere que la
adaptación al estrés térmico es un fenómeno complejo que involucra tanto la constitución genética como la capacidad de respuesta
a intervenciones nutricionales.
Análisis de semilla
por efecto de genotipo y tratamiento de fertilización foliar
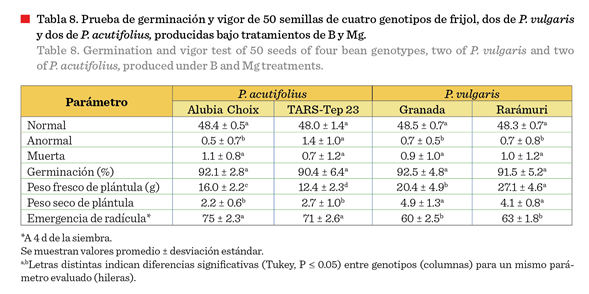
No hubo diferencias
significativas (P ≤ 0.05) entre genotipos para la variable de germinación,
resultando todos los materiales con germinación superior al 90 % (Tabla 8). La
prueba de vigor (emergencia de radícula a los 4 d) mostró diferencias significativas (P ≤ 0.05) entre genotipos, resultando superiores los genotipos de P. acutifolius
en comparación con P. vulgaris, desde un 10 % más de raíces emergidas en 4
d. Este resultado se debe probablemente a una cubierta de la testa de menor
grosor que la de P. vulgaris, tal y como lo indican Jiménez-Galindo y
Acosta-Gallegos (2012), quienes mencionaron que en algunos materiales, la germinación se genera a partir
del segundo día después de que se presenten las condiciones favorables para la
germinación por su rápida absorción de agua.
Prueba de germinación
y vigor de semilla por efecto de tratamiento
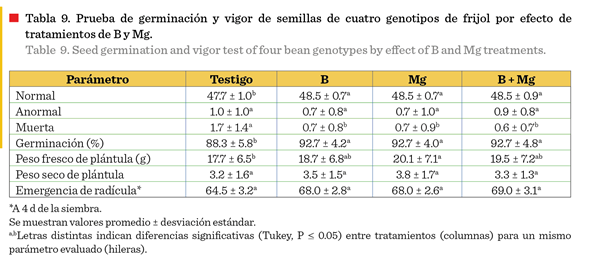
Los tratamientos de
fertilización foliar afectaron significativamente la calidad de semilla
cosechada. Los tratamientos B, Mg y B + Mg alcanzaron valores de germinación
superiores al 92 %, mientras que el testigo obtuvo un 88.3 % (Tabla 9). La
emergencia de radícula se incrementó de 64.5 % en el testigo hasta 68 % o 69 % en los tratamientos con fertilización
foliar. Hubo diferencias significativas en el número de semillas muertas, con
respecto al testigo, que presentó un mayor número, así como un menor porcentaje
de plántulas normales, lo que tuvo como resultado un porcentaje más bajo de
germinación. Esto indica que, la semilla tratada con fertilización foliar usando B y Mg fue superior significativamente en calidad
a la del testigo (Tabla 9), lo que permite recomendar este tratamiento para
elevar su calidad. Los tratamientos con B y Mg mejoraron significativamente la
viabilidad de la semilla, posiblemente a través de su papel en el metabolismo
de carbohidratos y la síntesis de proteínas
durante el desarrollo del embrión, como se observa en los valores
específicos de germinación y vigor (Tabla 9).
El efecto positivo de
la fertilización foliar sobre la calidad de semilla puede explicarse por varios mecanismos fisiológicos relacionados con
la nutrición de la planta madre durante el llenado de grano. En la mayoría de
las especies vegetales cuando sus semillas son sometidas a temperaturas mayores
a 30 °C se afecta de manera directa la producción de reguladores de crecimiento
como citoquininas y auxinas
(Chaves-Barrantes y Gutiérrez-Soto, 2017). Durante la etapa reproductiva
para obtener las semillas utilizadas para la prueba se presentaron en campo
temperaturas ≥ 30 °C. Si bien, las semillas se encuentran protegidas de manera
natural dentro de las vainas, los resultados obtenidos mostraron mayor cantidad
de semillas muertas en el tratamiento
testigo, lo que conlleva a una menor
germinación en comparación con los tratamientos donde la semilla se produjo
bajo aplicación de nutrición foliar con B y Mg. En el caso del tratamiento
testigo sin la fertilización foliar, se incrementó la presencia de semillas
muertas y un menor porcentaje de germinación pudo deberse a la deshidratación de los tejidos del embrión que provocan las altas
temperaturas en la formación de la semilla (Chaves-Barrantes y
Gutiérrez-Soto, 2017).
Además, el peso fresco
de la plántula del testigo fue inferior a todos los tratamientos con
fertilización foliar, debido a que las altas temperaturas afectan
considerablemente la permeabilidad de las membranas celulares en las radículas
(Kurepin y col., 2015). Esta respuesta positiva de la calidad de semilla a la
nutrición foliar tiene implicaciones importantes para los programas de
producción de semilla de frijol, especialmente en regiones donde las altas
temperaturas durante el llenado de grano pueden comprometer la viabilidad.
Los hallazgos
obtenidos ofrecen alternativas viables para sistemas productivos en regiones
propensas a altas temperaturas en el rango de las que se presentaron en la
región donde se llevó a cabo el estudio, donde la combinación de germoplasma
adaptado y manejo nutricional adecuado puede constituir una estrategia integral
para mantener la producción de frijol ante
condiciones ambientales cada vez más extremas. Sin embargo, es
importante reconocer que una limitación del presente estudio es la ausencia de
un diseño experimental que incluya diferentes niveles controlados de
temperatura, lo cual dificulta atribuir los efectos observados exclusivamente
al estrés térmico. No obstante, los resultados obtenidos reflejan la respuesta
de los genotipos y tratamientos bajo las condiciones térmicas naturales
prevalecientes durante los años de evaluación, por lo que futuras
investigaciones deberían considerar, el uso de ambientes controlados o
contrastantes para confirmar los mecanismos específicos de tolerancia al estrés
por temperatura.
CONCLUSIONES
La
fertilización foliar con micronutrientes demostró
ser una estrategia efectiva para contrarrestar los efectos adversos de las
altas temperaturas en el cultivo de frijol, evidenciando mecanismos
diferenciales de respuesta entre especies y variedades. Este estudio revela que
la aplicación de B y Mg, particularmente este último, no solo mejora parámetros
productivos, sino que también influye positivamente en la calidad fisiológica
de la semilla, aspecto fundamental para mantener la viabilidad del material genético
en condiciones de estrés térmico. La respuesta superior de los genotipos de P. vulgaris
sobre los de P. acutifolius en condiciones de altas temperaturas
contradice la percepción tradicional que señala al frijol Tepari como más
tolerante al calor. Este hallazgo sugiere que la adaptación al estrés térmico
es un fenómeno complejo que involucra tanto la constitución genética como la
capacidad de respuesta a intervenciones nutricionales.
AGRADECIMIENTOS
Los autores agradecen
al Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias
(INIFAP) por el financiamiento otorgado a través del proyecto "Desarrollo
de variedades mejoradas de frijol para
México" (SIGI: 16215236227). El
primer autor agradece al Consejo Nacional de Humanidades, Ciencias y
Tecnologías (Conahcyt) por la beca nacional otorgada para realizar estudios de
Maestría en Ciencias en Producción y Tecnología de Semillas, así como al
Tecnológico Nacional de México-Instituto Tecnológico de Roque, por el apoyo
brindado durante sus estudios de posgrado, del cual este trabajo forma parte de
su tesis.
Declaración de conflicto de
intereses
Los autores declararon
no tener conflictos de intereses de ningún tipo.
Referencias
Ahmad, W., Niaz, A., Kanwal, S., &
Rasheed, K. (2009). Role of boron in
plant growth: A review. Journal of Agricultural Research,
47(3), 329-338.
Anaya-López, J. L., Rojas-Tovar, L. M.,
Cisneros-López, H. C. y Acosta-Gallegos, J. A. (2022).
Rendimiento e índice de cosecha de germoplasma de frijol pinto y flor de mayo
bajo estrés por temperatura alta. Revista Fitotecnia Mexicana, 45(1),
33-33.
Baloch, F. S., Nadeem, M. A., Sönmez,
F., Habyarimana, E., Mustafa, Z., Karaköy, T., Cömertpay, G., Alsaleh, A.,
Çiftçi, V., Sun, S., Chung, G., & Chung, Y. S. (2022). Magnesium- a Forgotten Element: Phenotypic
Variation and Genome Wide Association Study in Turkish Common Bean Germplasm. Frontiers in Genetics,
13, 848663. https://doi.org/10.3389/fgene.2022.848663
Barrios-Gómez, E. J. y López-Castañeda,
C. (2009). Temperatura base y tasa de extensión foliar en frijol. Agrociencia, 43(1), 29-35. http://www.scielo.org.mx/scielo.php?script=sci_arttext&pid=S1405-31952009000100004
Beebe, S. E., Rao, I. M., Cajiao, C., & Grajales, M.
(2008). Selection for drought tolerance in common bean also improves yield in
phosphorus limited environments. Crop Science, 48, 582-592
Behera, B., Kancheti, M., Raza, M. B., Shiv, A., Mangal, V.,
Rathod, G., Altaf, M. A., Kumar, A., Aftab, T., Kumar, R., Tiwari, R. K., Lal, M. K., & Singh, B. (2023). Mechanistic insight on boron-mediated toxicity in plant vis-a-vis its
mitigation strategies: a review. International Journal of Phytoremediation,
25(1), 9-26. https://doi.org/10.1080/15226514.2022.2049694
Bornowski, N., Hart, J. P., Palacios, A.
V., Ogg, B., Brick, M. A., Hamilton, J. P., Beaver, J. S., Buell, C. R., &
Porch, T. (2023). Genetic variation in a tepary bean (Phaseolus acutifolius A.
Gray) diversity panel reveals loci associated with biotic stress resistance. The
Plant Genome, 16(3), e20363. https://doi.org/10.1002/tpg2.20363
Cakmak, I. y Yazici, A. (2010). Magnesio: El elemento
olvidado en la producción de cultivos. Informaciones Agronómicas-IPNI,
94(2), 23-25. https://www.ipni.net/ppiweb/iaecu.nsf/$file/2.+Magnesio.+El+elemento+olvidado.pdf
Chaves-Barrantes, N. F. y Gutiérrez-Soto, M. V. (2017).
Respuestas al estrés por calor en los cultivos. I. aspectos moleculares,
bioquímicos y fisiológicos. Agronomía
Mesoamericana, 28(1), 237-253. https://doi.org/10.15517/am.v28i1.21903
Cohen, J. (1988). Statistical power analysis for the
behavioral sciences (Second edition). Lawrence Erlbaum Associates.
Feng, Y., Fan, X., Suo, D., Zhang, S., Ma, Y., Wang, H.,
Guan, X., Yang, H., & Wang, C. (2023). Screening
of heat stress-regulating active fractions in mung beans. Frontiers
in Nutrition, 9, 1102752. https://doi.org/10.3389/fnut.2022.1102752.
Jiménez-Galindo, J. C. y Acosta-Gallegos,
J. A. (2012). Caracterización de genotipos criollos de frijol Tepari (Phaseolus
acutifolius A. Gray) y común (Phaseolus vulgaris L.) bajo temporal. Revista
Mexicana de Ciencias Agrícolas, 3(8), 1565-1577. http://www.scielo.org.mx/scielo.php?script=sci_arttext&pid=S2007-09342012000800007
Kirkby, E. & Rómheld, V. (2008). Micronutrientes en la
fisiología de las plantas: funciones, absorción y movilidad. Informaciones
Agronómicas, 68(2), 9-13. https://exa.unne.edu.ar/biologia/fisiologia.vegetal/MicronutrientesenlaFisiologia.pdf
Klaedtke, S. M., Cajiao, C., Grajales,
M., Polanía, J., Borrero, G., Guerrero, A., Rivera, M., Rao, I., Beebe, S. E.,
& Léon, J. (2012). Photosynthate remobilization capacity from
drought-adapted common bean (Phaseolus vulgaris L.) lines can improve yield
potential of interspecific populations within the secondary gene pool. Journal of Plant Breeding and Crop Science, 4(4), 49-61. https://doi.org/10.5897/JPBCS11.087
Kurepin, L. V., Park, J. M., &
Lazarovits, G. (2015). Burkholderia phytofirmans-induced shoot and root growth
promotion is associated with endogenous changes
in plant growth hormone levels. Plant
Growth Regulation, 75(1), 199-207. https://doi.org/10.1007/s10725-014-9944-6
Mora, M., Ordaz, V., Castellanos, J. Z., Santelises, A. A.,
Gavi, F. y Volke, V. (2001). Sistemas de labranza y sus efectos en algunas
propiedades físicas en un vertisol, después de cuatro años de manejo. Terra
Latinoamericana, 19(1), 67-74.
Omae, H., Kumar, A., & Shono, M. (2012). Adaptation to
high temperature and water deficit in the common bean (Phaseolus vulgaris L.)
during the reproductive period. Journal of Botany, 2012, 803413. https://doi.org/10.1155/2012/803413
Polania, J., Rao, I. M., Cajiao, C., Rivera, M., Raatz, B.,
& Beebe, S. (2016). Physiological traits associated with
drought resistance in Andean and Mesoamerican genotypes of common bean (Phaseolus
vulgaris L.). Euphytica, 210(1), 17-29. https://doi.org/10.1007/s10681-016-1691-5
Porch, T. G. & Jahn, M. (2001).
Effects of high-temperature stress on microsporogenesis in heat-sensitive and
heat-tolerant genotypes of Phaseolus vulgaris. Plant, Cell and
Environment, 24(7), 723-731. https://doi.org/10.1046/j.1365-3040.2001.00716.x
Rainey, K. M. y Griffiths, P. D. (2005) Respuesta diferencial
de genotipos de frijol común a altas temperaturas. Revista de la Sociedad
Americana de Ciencias Hortícolas, 130, 18-23. https://doi.org/10.21273/JASHS.130.1.18
Siddiqui, M. H., Alamri, S. A., Al-Khaishany, M. Y. Y.,
Al-Qutami, M. A., Ali, H. M., Al-Whaibi, M. H., Al-Wahibi, M. S., &
Alharby, H. F. (2018). Mitigation of adverse effects of heat
stress on Vicia faba by exogenous
application of magnesium. Saudi Journal of Biological
Sciences, 25(7), 1393-1401.
Stagnari, F., Onofri, A., & Pisante,
M. (2009). Response of French Bean (Phaseolus vulgaris L.) Cultivars to Foliar Applications of Magnesium. Italian
Journal of Agronomy, 3, 101-110. https://doi.org/10.4081/ija.2009.101
Tapia, G., Méndez, J., Inostroza, L.,
& Lozano, C. (2022). Water Shortage
Affects Vegetative and Reproductive Stages of Common Bean (Phaseolus
vulgaris) Chilean Landraces, Differentially Impacting Grain Yield
Components. Plants (Basel, Switzerland), 11(6), 749. https://doi.org/10.3390/plants11060749
Tene, T. M., Sari, H., Canci, H.,
Maaruf, A., Eker, T., & Toker, C. (2023). Traits Related to Heat Stress in
Phaseolus Species. Agriculture, 13(5), 953.
https://doi.org/10.3390/agriculture13050953