https://doi.org/10.29059/cienciauat.v20i2.2052
Estructuras G-cuádruples en
genomas virales: prometedoras dianas moleculares para fines terapéuticos
G-quadruplex structures in viral genomes:
Promising molecular targets for therapeutic purposes
Estructuras
G-cuádruples en genomas virales
Joseline Ontiveros-Euán, Stefano
Decle-Carrasco, Nuvia Kantún-Moreno*
*Correspondencia:
nuvia.kantun@correo.uady.mx/Fecha de recepción: 26 de agosto de 2025/Fecha de
aceptación: 16 de enero de 2026/Fecha de publicación: 29 de enero de 2026.
Universidad Autónoma de Yucatán, Centro
de Investigaciones Regionales, Dr. Hideyo Noguchi, Laboratorio de Virología,
calle 43 núm. 613 x calle 90, col. Inalámbrica, Mérida, Yucatán, México, C. P.
97069.
Resumen
Las
G-cuádruples (G4s) son estructuras secundarias de ácidos nucleicos presentes en
todos los reinos biológicos. En humanos, han sido identificadas principalmente
en los telómeros y en los promotores de oncogenes. En virus, las G4s han sido
evidenciadas en genomas de ADN y ARN, aunque su relevancia funcional en los
procesos infectivos y de patogénesis permanece escasamente caracterizada. El
objetivo del presente trabajo fue dar a conocer
el estado del arte actual de las estructuras G4s en virus, abarcando su
identificación, características estructurales, interactores (proteínas virales
y celulares), y su implicación en la regulación del ciclo viral. La evidencia
actual demuestra que las estructuras G4s están presentes en todos los grupos de
la clasificación de Baltimore y participan en la regulación de procesos
esenciales, como la replicación, transcripción y traducción de proteínas. Los
hallazgos científicos posicionan a las G4s
como prometedoras dianas moleculares con miras a ser consideradas como
una opción innovadora y viable para fines terapéuticos.
PALABRAS CLAVE: G-cuádruples, virus de humanos, genomas, antivirales, dianas moleculares.
ABSTRACT
G-quadruplexes (G4s) are secondary structures of nucleic acids present
in all biological kingdoms. In humans, they have been identified mainly in telomeres and oncogene promoters. In viruses, G4s have
been found in DNA and RNA genomes, although their functional relevance in
infection and pathogenesis processes remains poorly characterized. The aim of
this paper is to present the current state of the art of G4s structures in
viruses, including their identification, structural features, interactors
(viral and cellular proteins), and their involvement in the viral cycle
regulation. Current evidence shows that G4s structures are present in all
Baltimore classification groups and participate in the regulation of essential
processes, such as replication, transcription, and protein translation. These
scientific findings position G4s as promising molecular targets with a view to
being considered as an innovative and viable option for therapeutic purposes.
KEYWORDS: G-quadruplexes, human viruses, genomes, antivirals, molecular
targets.

INTRODUCCIÓN
Los virus son agentes infecciosos
submicroscópicos que dependen de organismos huéspedes para replicarse. Su
estructura básica consiste en material genético, ya sea ácido
desoxirribonucleico (ADN) o ácido ribonucleico (ARN) encapsulado en una
cubierta proteica (Ruggiero y Richter,
2020). Estos parásitos moleculares, considerados las entidades
biológicas más abundantes y diversas de la tierra, infectan a todos los tipos de organismos, desde bacterias y hongos hasta plantas y animales, incluidos los humanos, evolucionando
simultáneamente con la vida misma (Koonin y
col., 2020). Se caracterizan en
términos de su morfología, tamaño y naturaleza del genoma, pero al no
replicarse por sí mismos, necesitan de la maquinaria biosintética celular de
otros organismos para propagarse; en consecuencia, infectan al hospedero, y con
ello, son los responsables de brotes, epidemias y pandemias que afectan a la
salud humana y otras especies (Edwards y col., 2022).
En la actualidad, se han logrado
avances significativos en la identificación de dianas terapéuticas y desarrollo
de antivirales específicos contra algunos virus humanos. Sin embargo, aún
existen desafíos de salud pública debido a la falta de vacunas o tratamientos
eficaces contra diversos virus humanos (Hargrave y col., 2021). Un caso
emblemático es el del virus de la inmunodeficiencia humana (VIH), para el cual
existen antirretrovirales que controlan la infección, pero aún no se dispone de
una vacuna aprobada. Esta necesidad se ve acentuada por la rápida evolución
viral, que exige reevaluar fármacos
existentes (reposicionamiento) e investigar
nuevas dianas moleculares (Yan y col., 2023). Precisamente, en dicho
punto, las estructuras no canónicas de ácidos
nucleicos (aquellas diferentes a la doble hélice) como las G-cuádruples,
han sido de interés científico ante la extraordinaria diversidad de los genomas
virales. Estas se pueden encontrar en forma monocatenaria o bicatenaria de ADN
y ARN, empaquetados en segmentos o en una
sola pieza y de manera lineal, segmentado
o circular (Koonin y col., 2022).
El ADN es una macromolécula de mucha
importancia biológica, puesto que contiene la información genética que hace
únicos a los organismos vivos, y a los virus. Además, ésta es la base para la
construcción del ARN. Cabe destacar que el ADN es una molécula altamente
polimórfica o con una elevada flexibilidad conformacional (Pont y
González-García, 2022). El ADN-b es la forma estructural más común en las células, la cual fue descrita por
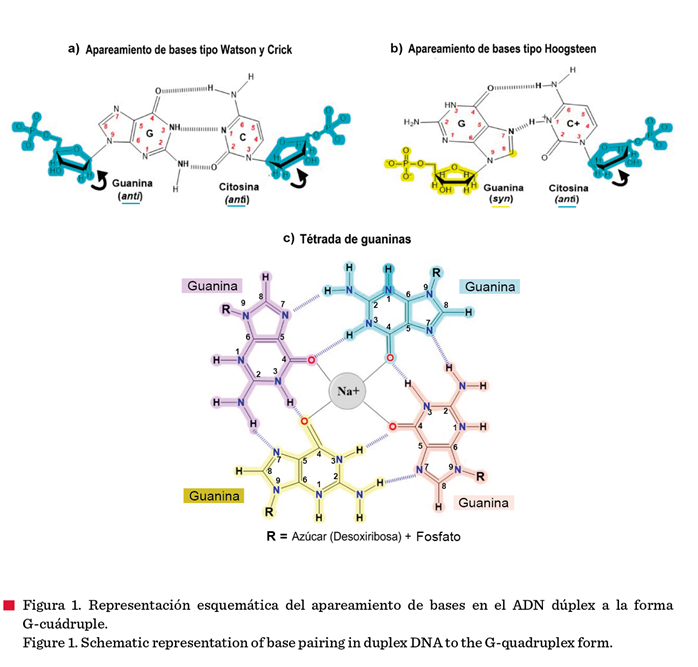
Watson y Crick en 1958 (Figura 1a). Sin embargo, este puede ser susceptible a
estrés torsional que puede dar lugar a cambios conformacionales por la
presencia de regiones ricas en guaninas (G) y citocinas (C), alteraciones en la concentración intracelular de iones,
modificaciones de las bases (mutaciones) y la presencia de ciertos fármacos,
entre otros. Todos esos factores pueden causar que el ADN-b adopte una
conformación diferente. A dichas estructuras se les reconoce como “formas no canónicas del ADN”. Una de estas
estructuras de importancia actual son las G-cuádruples (G4s) (Teng y col.,
2021; Pont y González-García, 2022; Sato y Knipscheer, 2023).
Las G4s representan interesantes dianas
de estudio, ya que se forman en el ADN o ARN, en específico, en regiones ricas
en G. Abundan en los organismos vivos, e incluso en los virus, por lo que su
localización en el genoma y su función son de interés científico. El ADN-G4 fue
presentado por primera vez por Gellert y col. (1962), pero no fue hasta hace
algunos años que estas estructuras llamaron la atención de la comunidad
científica debido a su relación con diversos procesos biológicos como la
transcripción, traducción, replicación del ADN y en la inestabilidad genómica
de genes, y por su potencial en aplicaciones terapéuticas como dianas
moleculares para el cáncer, sistemas de administración de fármacos y de
enfermedades, y como antivirales (Abiri y col., 2021; Artusi y col., 2021;
Datta y col., 2021; Debbarma y Acharya, 2022).
Los primeros estudios consistieron en
la identificación de estas estructuras tetracatenarias en el genoma humano, luego en el diseño de ligandos específicos
para la búsqueda de medicamentos contra el cáncer, y no mucho tiempo después se
mapearon mediante el uso de técnicas
moleculares en el genoma de algunos virus, en donde todavía se están elucidando
sus funciones biológicas (Ruggiero y col., 2021).
Diferentes estudios han evidenciado la
presencia de estas estructuras en células vivas. Summers y col. (2021) emplearon anticuerpos específicos para
visualizar G4s teloméricos en Stylonychia lemnae y anticuerpos de alta
afinidad para G4s de ADN y ARN en células de mamíferos por medio de
inmunofluorescencia. Recientemente, diseñaron sondas para el estudio in vivo
de la dinámica molecular de las G4s en
tiempo real, utilizando DAOTA-M2, una
sonda fluorescente que identifica G4s en
células vivas, para estudiar dichas estructuras y sus interacciones con
aglutinantes en células epiteliales del
osteosarcoma óseo humano (U2OS). Otro ejemplo, es el trabajo de Di-Antonio y col. (2020) con la sonda fluorescente
específica SiR-PyPDS, la cual combina el fluoróforo rojo de silicón-rodamina
(SiR) con un análogo del ligando G4 piridostatina (PyPDS). Estas nuevas
metodologías podrían ser de utilidad para estudiar su función y dinámica ante infecciones virales.
En este contexto, la investigación de
G4s en virus humanos es una opción muy atractiva de estudio, puesto a que se le
ha relacionado con la replicación y la recombinación viral, en la regulación de
la virulencia a través de la expresión génica y en pasos claves en el ciclo
infectivo que es muy particular de cada virus estudiado.
El objetivo del
presente trabajo fue dar a conocer el estado del arte actual de las estructuras
G4s en genomas virales, abarcando su identificación, características
estructurales intrínsecas, interactores como proteínas virales y celulares, y
su implicación en la regulación del ciclo viral.
Procedimiento de
búsqueda de información
La búsqueda bibliográfica se realizó en
la base de datos PubMed, desarrollada por el Centro Nacional de Información
Biotecnológica (NCBI, por sus siglas en inglés: National Center for Biotechnology Information) de los Estados Unidos.
La estrategia consistió en el empleo de términos clave en inglés: “estructura y
función de las G-cuádruples” (G-quadruplex structural and functions),
“G-cuádruples en virus" (G-quadruplex in virus), “actividad antiviral de
G-cuádruples” (antiviral activity of G-quadruplex) y “aplicaciones de las
G-cuádruples” (applications of G-quadruplex). Se incluyeron tanto artículos
originales como revisiones científicas publicadas en los últimos 15 años y
algunas referencias clásicas y relevantes en
virus, dando mayor énfasis a aquellos trabajos publicados en los últimos
5 años. Posteriormente, se clasificó y revisó la información, destacando en este trabajo sus aspectos estructurales y los
estudios actuales en virus de importancia médica, considerándose su unción
demostrada.
Aspectos estructurales de las G4s
Las G4s fueron propuestas por primera
vez por Gellert y col. (1962), quienes publicaron la primera estructura de un
tetrámero planar de G (G-tétradas) formando
un gel; posteriormente, Henderson y col. (1987) caracterizaron la
estructura completa de una ADN-G4 en los telómeros humanos. Este hallazgo fue
el inicio de múltiples investigaciones para identificarlas en el genoma humano (incluyendo promotores de oncogenes), sus funciones
biológicas y posibles aplicaciones, debido a que, su localización no es aleatoria;
más adelante, los estudios se enfocaron en otros organismos, incluyendo virus.
A partir del descubrimiento de G4 en papovirus SV-40 (Tuesuwan y col., 2008),
las investigaciones han ido en aumento, asociadas al creciente interés en la
elucidación de estas estructuras en genomas virales, la interacción de las G4s
con las proteínas del hospedero y su función en células infectadas.
Las G4s son estructuras de ácido
nucleico no canónicas que son ubicuas y que pueden formarse en secuencias ricas
en G del ADN y ARN. Se conforman por más de dos unidades planas cuadradas,
denominadas G-tétradas, que están apiladas una sobre otra. Cada tétrada se
forma con cuatro G que se mantienen en una disposición plana a través de
puentes de hidrógeno de tipo Hoogsteen (Figura 1b) con un catión monovalente
central, preferentemente K+ o Na+ (Figura 1c); la
presencia de dichos iones estabiliza a las G4s. Dependiendo del tamaño del
catión, este puede situarse en el mismo plano
de la G-tétrada, formando 4 enlaces covalentes coordinados con los
átomos de oxígeno de las G4s (en el caso del Na+, Figura 1c) u 8
enlaces con el K+, localizándose entre dos tétradas en un arreglo bipiramidal
anti prismático (Pont y González-García, 2022).
Las G4s se forman a partir de
secuencias con el motivo
G2−5-NL1G2-5NL2G2-5NL3G2-5, de las que de 2 G a 5 G son separadas por
horquillas (en inglés, loops) de hasta
7 nucleótidos mixtos (Puig-Lombardi y Londoño-Vallejo, 2020). Sin
embargo, esta definición ha sido ampliada para integrar G4s con imperfecciones,
como lo son protuberancias (bulges) y apareamientos erróneos (mistmatches),
que resultan de la presencia de bases distintas a la G en una o más G-tétradas,
diferenciándose del motivo G4 canónico (Das y col., 2021). La existencia de G4s
atípicas ha sido validada mediante experimentos in vitro e in vivo
y actualmente se consideran en la
identificación in silico con programas
bioinformáticos (Puig-Lombardi y Londoño-Vallejo, 2020).
Las estructuras G4s pueden adoptar
múltiples conformaciones en los ácidos nucleicos (Pont y González-García,
2022). Al ser polimórficas, su topología se basa en el tipo de loop, la
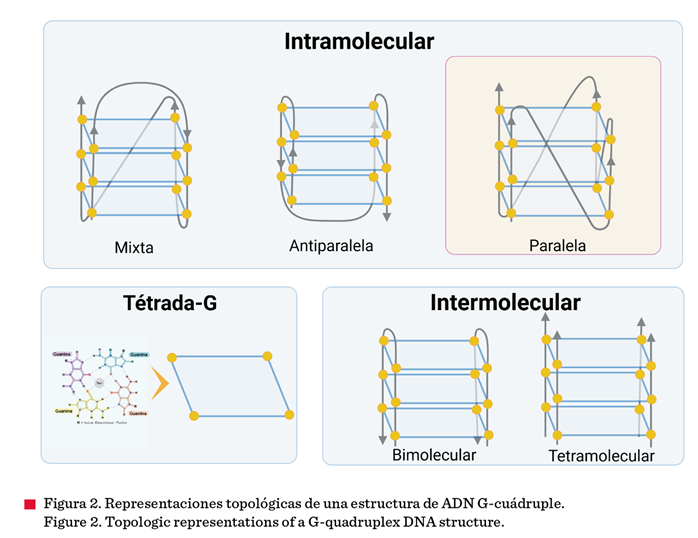
orientación y el número de hebras. En este contexto, se clasifican en G4s
intramoleculares o intermoleculares. El primer tipo se forma a partir de una
hebra y el segundo tipo contiene más de dos hebras que pueden ser G4s
bimoleculares (dos hebras ricas en G) o tetramoleculares (cuatro hebras)
(Figura 2). De acuerdo con la orientación de las hebras pueden adoptar
conformaciones de tipo: paralela, antiparalela o mixta (Figura 2). En las G4s paralelas, todas las hebras están orientadas
en la misma dirección (4 + 0) mientras que, en las conformaciones
antiparalelas, la orientación es opuesta para la mitad del total de las hebras
(2 + 2). Por su parte, las G4s mixtas o híbridas tienen una hebra en dirección opuesta a las tres restantes (3 + 1)
(Biver, 2022). De igual manera, estas estructuras se conectan por diferentes
tipos de loops; el tipo hélice conecta las hebras paralelas, la lateral
puede conectar hebras antiparalelas adyacentes y la diagonal une hebras
antiparalelas a través del núcleo de la G-tétrada.
En el caso de las G4s de ARN, son
estructuras mucho más estables termodinámicamente en comparación con las de ADN
(Fay y col., 2017). La presencia del grupo 2'-hidroxilo en la ribosa del ARN
posibilita más interacciones intramoleculares dentro de la misma molécula,
formándose enlaces de hidrógeno extras, favoreciendo así a la hidratación molecular y a su estabilidad (Caterino
y Paeschke, 2022). Estas mismas propiedades biofísicas promueven que las G4s
sean menos polimórficas en el ARN. Se considera que la conformación G4 paralela
es la más predominante y se debe a la
conformación anti (Figura 2) de los enlaces glicosídicos en los ribonucleótidos del
ARN (Lyu y col., 2021).
La formación de estas estructuras
depende de la composición de los nucleótidos de las hebras y de otros factores
externos como la fuerza iónica o la temperatura. Es importante señalar que, a
la hora de evaluar el potencial de la formación de las G4s y sus posibles
funciones, es necesario tener en cuenta sus propiedades termodinámicas y
cinéticas. Ellas pueden interactuar específicamente con proteínas o incluso con
ligandos más pequeños, que pueden competir con las proteínas o imitar algunas
de sus funciones (Caterino y Paeschke, 2022).
Estructuras G4s en virus
Los virus se pueden clasificar por
diferentes características, como la estructura de la cápside, tipo de ácido
nucleico y polaridad, presencia de envoltura o tipo de infección. La taxonomía
de los virus es dada por el Comité Internacional de Taxonomía de Virus (ICTV)
(https://ictv.global/taxonomy/) y toma en consideración
la clasificación de Baltimore, la cual se basa en el tipo de genoma
viral y la ruta por el cual su información genética se transfiere (Ruggiero y
Richter, 2020). Se categorizan en 7 grupos o clases; grupo 1: ADN bicatenario
(ADNbc); grupo 2: ADN monocatenario (ADNmc);
grupo 3: ARN bicatenario (ARNbc); grupo 4: ARN monocatenario positivo (ARNmc+);
grupo 5: ARN monocatenario negativo (ARNmc-); grupo 6: ARNmc
retrotranscrito (ARNmcRT) y grupo 7: ADNbc retrotranscrito (ADNbcRT) (Koonin y
col., 2021).
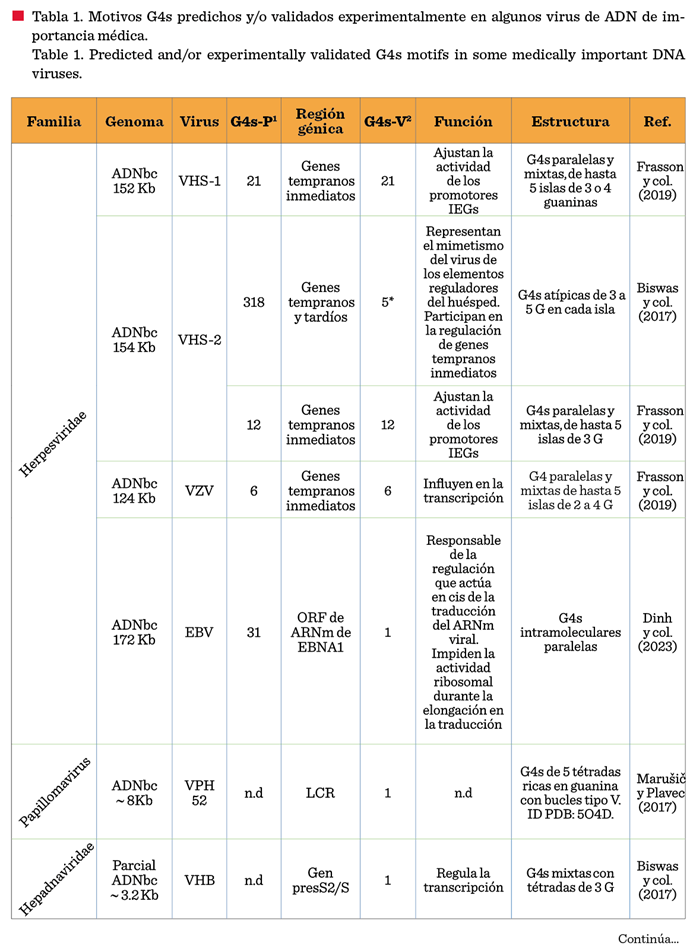
Se ha descrito la presencia de G4s en
la mayoría de los grupos de Baltimore, evidenciando que son una característica
de cada familia viral (Lavezzo y col., 2018). Los primeros estudios
identificaron G4s en genomas virales de importancia médica mediante programas
bioinformáticos, seguidos de validaciones in vitro. Sin embargo, pocos
trabajos han confirmado experimentalmente las G4s predichas en virus humanos y
su función biológica. Revisiones recientes destacan algunas interacciones entre
las G4s y sus funciones en virus (Lavezzo y col., 2018; Abiri y col., 2021;
Zareie y col., 2024).
La alta densidad y ubicación de las G4s
en los genomas virales sugieren un papel clave en la replicación,
recombinación, regulación de la virulencia y expresión génica (Abiri y col.,
2021; Zareie y col., 2024). Por ejemplo, en el virus del herpes simple 1
(VHS-1), su genoma rico en Guanina-Citosina (GC) contiene secuencias repetidas
que forman G4s estables, favoreciendo la replicación del ADN viral (Frasson y
col., 2019). En el virus de la inmunodeficiencia
humana (VIH-1), el promotor LTR presenta una región rica en G conservadas en la
región U3, generando G4s funcionales que regulan la replicación y
transcripción viral. Debido a su relevancia, las G4 son consideradas blancos
terapéuticos en virus de ADN y ARN como Zika, Virus de la hepatitis B (VHB),
Ébola, Herpesvirus, SARS-CoV-2 y Nipah
(Ruggiero y Richter, 2020). A continuación, se presentan ejemplos de su
caracterización en virus patógenos humanos, clasificados según su composición
genómica.
G4s en virus de ADN
Entre los virus
ADNbc, la familia Herperviridae presenta una alta frecuencia de motivos G4s en regiones
reguladoras y repetidas del genoma (Ruggiero y Richter, 2020). Los VHS-1 y
VHS-2 son los que presentan una mayor cantidad
de G4s detectadas hasta el momento.
Muchas de estas se encuentran en los
genes de expresión inmediata temprana (IEGs,
por sus siglas en inglés: Inmediate Early Gene), sugiriendo que actúan
como moduladores de la transcripción a nivel del promotor (Frasson y col.,
2019). Por su parte, se han identificado 36
motivos G4s dispersos en todo el genoma de los citomegalovirus, 20 de
ellos entre los genes inmediatos, tempranos y tardíos. Estos, en su mayoría,
adoptando una conformación G4 paralela y actuando en el control de la expresión génica viral, por lo que se
especula que participan en el establecimiento de la infección latente en esos
virus (Ravichandran y col., 2018).
También, se ha demostrado la presencia
de estructuras G4s conservadas en muchas de las especies de la familia
Adenoviridae, pero con mayor porcentaje en cepas que infectan al humano. Cabe
destacar, el papel de estos virus en la biotecnología roja (centrada en la
medicina y salud humana), por su papel como vectores de transferencia en
terapia génica. La caracterización funcional de dichas estructuras evidenció la
presencia de 15 G4s conservadas en varias cepas de adenovirus, las cuales están
situadas en genes esenciales (E1B, E2B, L3, entre otros) para su ciclo
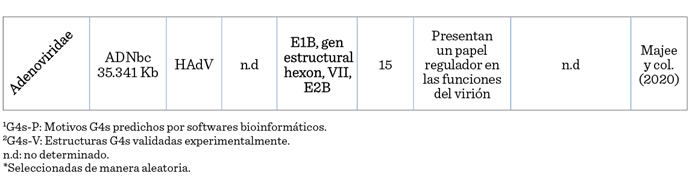
infectivo (Bohálová y col., 2021). La estabilización de los motivos G4s en
estos virus permiten la producción de viriones (Majee y col., 2020). Varios
estudios apoyan firmemente la existencia de
estructuras G4s en el genoma de los adenovirus humanos (HAdV) que
afectan a la multiplicación del virus y plantean
que dichas estructuras pueden influir en la eficacia de los vectores de
liberación del gen o incluso en el ciclo de vida de los HAdV (Majee y col.,
2020).
En los papilomavirus humanos, se ha
detectado la presencia de G4s en solo 8 genotipos, 2 de ellos son de alto
riesgo oncogénico. Estas estructuras se encontraron en la región larga de
control (LCR, por sus siglas en inglés: Long Control Region) de los virus del
papiloma humano (VPH) 52 y 58 (Marušič y Plavec, 2019), en el gen L1 de VPH 57,
en el gen E1 de los VPH 32 y 42, y en el gen
E4/E2 de los VPH 3, 9 y 25; su posición sugiere que la formación de
estas estructuras en el genoma viral está relacionada con la transcripción y
pueden ser zonas de interés para el diseño de antivirales (Tlučková y col.,
2013). Por su parte, Marušič y col. (2017) evidenciaron el potencial de 8
secuencias ricas en G en los genomas de los VPH de alto riesgo 16, 18, 52 y 58.
También indicaron la formación de una G4 inestable en L2 de VPH 16, debido a
una alta cantidad de polimorfismos estructurales
que favorecen la competencia con la formación| de horquillas. Además,
determinaron que las G4s de los VPH 52 y 58 eran las más estables.
Existe poca
información de los virus de ADNmc;
sin embargo, se reportó la existencia de G4s
en virus adenoasociados y las han relacionado con nucleofosmina, una
proteína encargada de la estabilidad del genoma, reparación y replicación del
ADN. En dicho contexto, se reportó que su interacción con esta podría resultar en la inhibición de la
replicación del ADN viral y hasta celular (Satkunanathan y col., 2017).
El VHB es el único virus humano
perteneciente al grupo de ADNbcRT que presenta estructuras G4s en su genoma. Se
identificó una G4 conservada tipo mixta en el promotor preS2 del gen S, y se
observó que la estabilización de esta,
mediante ligandos, incrementa la transcripción
viral, lo que resultó en una mayor expresión del antígeno de superficie
(HBsAg) del virus de la hepatitis B y en una mayor secreción de viriones
(Biswas y col., 2017). También, se han identificado estructuras G4s en el
promotor preC del VHB mediante ensayos biofísicos y experimentos in vitro
en células HepG2 infectadas con este virus, reportándose un efecto inhibitorio
en la replicación viral. Dicha región promotora es esencial para la
transcripción del gen C y la posterior traducción de proteínas de la
nucleocápside (Meier-Stephenson y col., 2021). En la Tabla 1, se presentan
algunas G4s reportadas de distintos virus de ADN.
G4s en virus de ARN
Se ha reportado una mayor cantidad de
motivos de G4s en virus de ARNmc de hebra positiva y negativa (Fay y col.,
2017). Sin embargo, en los virus de ARNbc no hay información reportada y en los virus de ARNmcRT son pocas las G4s descritas (Ruggiero y Richter,
2020: Abiri y col., 2021).
Entre los virus ARNmc- se ha detectado
una G4 en el gen L del virus del ébola. Este gen codifica para la ARN
polimerasa dependiente del ARN viral, responsable de la replicación y
transcripción; la estabilización de esta G4 por medio de ligandos puede disminuir
la actividad de la polimerasa e inhibir la replicación viral (Wang y col.,
2016b). En el virus de la influenza H1N1 también se ha detectado, con mayor
frecuencia, posibles secuencias formadoras de G4 (SFG4) en el segmento NP del
genoma viral. Dicho gen codifica una nucleoproteína que se encarga de la
replicación viral y, por lo tanto, la
formación de esta G4 podría ser una posible diana molecular (Brázda y
col., 2023).
Existe una gran variedad de virus de
ARNmc+ en los que se ha reportado la presencia de G4s,
pertenecientes a familias como Flaviviridae y Coronaviridae
(Lavezzo y col., 2018; Ji y col., 2021; Cui y Zhang, 2020). En la familia Flaviviridae,
se descubrieron 4 G4s paralelas en el genoma del virus del Zika, 2 se
localizaron en la región codificante del gen NS5, y las otras 2, en los genes
NS2 y NS4B, dichos datos implican a estas estructuras en la patogénesis del
virus (Gemmill y col., 2024). En VHC se identificaron 6 SFG4, siendo la más
estable una G4 en el gen de la proteína C o de la cápside. Un estudio in
vivo reveló que el derivado de piridostatina (PDP), un ligando G4, puede
estabilizar esta G4 e inhibir la replicación intracelular y la traducción (Wang
y col., 2016a). Otro estudio demostró que la unión de la proteína celular,
nucleolina, con esta estructura G4 puede
frenar la replicación viral. Ambos, actuando como un posible mecanismo de inmunidad antiviral (Bian y col., 2019).
En los Coronoviridae, se han
detectado G4s en tres de los virus más patógenos en humanos, raros y altamente
contagiosos de dicha familia, responsables de emergencias sanitarias a nivel
mundial. En el virus SARS-CoV se identificó que el subdominio SUD-M de la
proteína no estructural multidominio 3 (Nsp3) interacciona con una G4 y este es
esencial para su actividad de
replicación/transcripción. También, el mismo subdominio, se ha
encontrado en el coronavirus del síndrome respiratorio de Oriente Medio
(Mers-CoV) (Kusov y col., 2015). Por medio de un análisis in silico, se
elucidaron G4s en la región codificante de la proteína S en 5 coronavirus, incluyendo al SARS-CoV, SARS-CoV-2 y MERS-CoV (Cui y Zhang, 2020). En el
SARS-CoV-2 se identificaron 25 (Ji y col., 2021).
Aunque no hay trabajos que describan la
existencia G4 en virus de ARNbc, Lavezzo y col. (2018) mencionaron que, en los
virus de ese grupo pudieron determinar pocas SFG4 potenciales, sin embargo, todas ellas se encontraban altamente
conservadas, sugiriendo un potencial interés biológico. Bohálová y col. (2021)
reportaron que los virus responsables de enfermedades agudas, como el rotavirus, un virus que pertenece a este
grupo, presenta frecuencias muy bajas de motivos de G4s (0.68 por 1000 nt).
El VIH, agente etiológico del síndrome
de inmunodeficiencia adquirida (SIDA) pertenece al grupo de los virus ARNmcRT.
En el genoma de este virus, se han reportado secuencias que pueden formar G4s
en la región gag, la cual codifica para formar sus proteínas estructurales del
virus (Piekna-Przybylska y col., 2014). Además, se han encontrado múltiples G4s
en la región U3, situada en el extremo 5' de la forma proviral del VIH-1,
superpuestas con los tres sitios de unión de Sp1 del promotor viral, en las formas
de ARN y ADNmc. Ello sugiere que, cuando ocurre la infección viral, las G4s se
pliegan en el genoma del ARN como en el promotor del provirus del VIH-1 y
tienen un papel importante en la regulación del virus (Piekna-Przybylska y
col., 2014; Amrane y col., 2022). Diversas G4s han sido localizadas en el
promotor LTR proviral, en esta región se pueden formar 3 estructuras
superpuestas: LTR-II, LTR III y LTR-IV. La región LTR III puede formar G4s
híbridas con una conformación cuádruplex-dúplex y la región LTR-IV presenta una
G4 paralela en el final 3´. La estabilización de la primera G4 suprime la
transcripción viral, y la segunda, realiza todo lo contrario, es decir,
incrementa la actividad transcripcional (De-Nicola y col., 2016; Butovskaya y
col., 2018). En la Tabla 2, se presenta
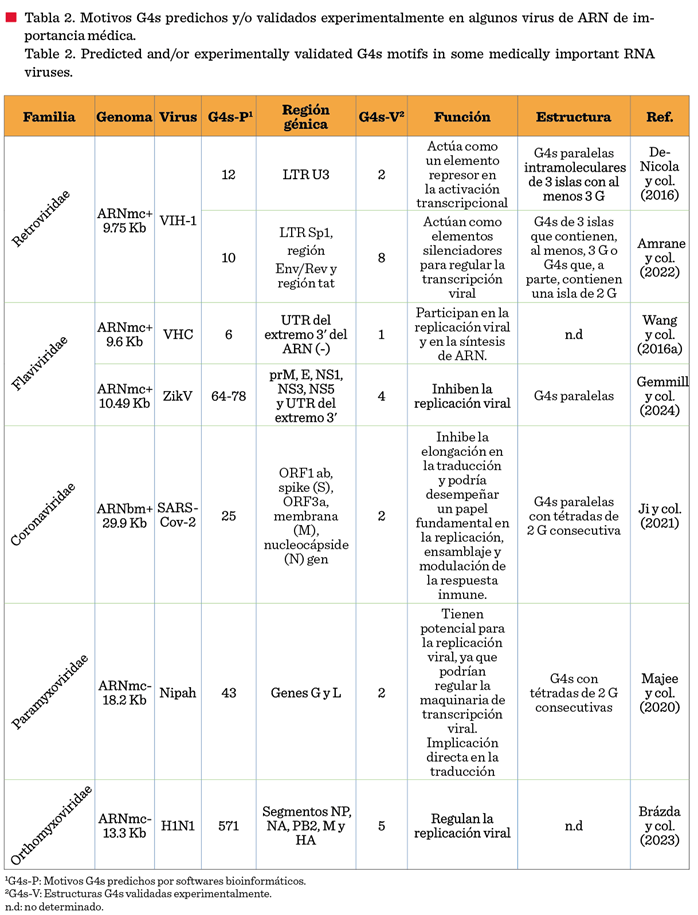
información de algunas G4s de virus de ARN.
La participación de las estructuras G4s
en procesos virales esenciales, las vuelve de interés como blancos moleculares
para el diseño de antivirales. Se han estudiado diversos ligandos,
particularmente moléculas de bajo peso molecular, de naturaleza predominantemente
aromática y planar, lo que les confiere que puedan intercalarse o apilarse
sobre las tétradas de G de las G4s (Abiri y col., 2021). El mecanismo de acción
más estudiado es la estabilización de estas estructuras, impidiendo que
proteínas celulares actúen sobre las G4s e interfiriendo así procesos como la
replicación, transcripción de genes o traducción de proteínas virales. En otras
palabras, se "bloquea" la estructura y se impide que enzimas
celulares esenciales para la replicación viral, como las helicasas (p. ej.,
DHX36, DHX9, FANCJ, PIF1) y polimerasas (tanto de ADN como de ARN), la
desenrollen. Sin embargo, también se propone
el empleo de ligandos para revertir la estructura G4 a una forma
monocatenaria y regular negativamente la persistencia viral (Zareie y col.,
2024).
Entre los ligandos de estructuras de
G4s más investigados por su potencial actividad antiviral se encuentran: 1) las
porfirinas y sus derivados, como TMPyP4 y N-metil mesoporfirina IX (NMM), que
se han evaluado frente a una amplia gama de virus, como el de la viruela símica
(MPXV), el virus Zika, el SARS-CoV-2, VHS-1 y VIH, no obstante, es importante
señalar que, algunas porfirinas también han sido propuestas como compuestos con
potencial carcinogénico; 2) compuestos de bisquinolinio, como PhenDC3, que han demostrado
eficacia contra el virus de Epstein-Barr (VEB), el de la hepatitis B (VHB), el
citomegalovirus humano (HCMV) y el VPH; 3) las acridinas, entre las que destaca
BRACO-19, ligando que ha demostrado capacidad para regular la expresión viral
por sí solo en diversos patógenos, como el VIH-1, MPXV, SARS-CoV-2 y VPH; 4)
PDS y sus derivados, que han sido activos
contra virus como el VPH, HCMV, VIH y
SARS-CoV-2 (Ruggiero y col., 2021; Zareie y col., 2024).
Un desafío crítico actual, en el
desarrollo de estas terapias, es la selectividad. En dicho contexto, se están
explorando estrategias terapéuticas alternativas y más selectivas en algunos
virus para evitar los efectos fuera de diana (por sus siglas en inglés: off-target effects), como la unión inespecífica a los G4s
del hospedero que podría desencadenar efectos adversos (p. ej., citotoxicidad).
Entre estas estrategias se incluyen el diseño de oligonucleótidos antisentido,
la estabilización de G4 mediada por el sistema CRISPR-Cas9, el empleo de ARN de
interferencia (ARNi) y el desarrollo de
aptámeros (Zareie y col., 2024).
CONCLUSIONES
El estudio de las
estructuras G4s en virus representa
un campo de investigación en constante crecimiento, con un enorme potencial en
biomedicina. Comprender su biología estructural y funcional no solo permite
desentrañar los mecanismos que regulan
procesos clave en el ciclo infectivo de los virus, sino que también abre la puerta al desarrollo de nuevas aplicaciones
en la virología. Se han propuesto con un doble propósito, ya sea como dianas
terapéuticas para el desarrollo de nuevos tratamientos o como herramientas
biotecnológicas en sistemas de diagnóstico de enfermedades y administración de
fármacos. Así, estas fascinantes estructuras
no canónicas de ácido nucleico están redefiniendo la manera en que se
entiende al ADN o ARN en la virología moderna.
Agradecimientos
Al proyecto Ciencia de frontera
CF-2023-I-600 “Evidenciando G-cuádruples en el virus del papiloma humano tipo
16: Una nueva visión estructural de su ADN como blanco terapéutico”, el cual es
financiado por SECIHTI, antes CONAHCYT.
Declaración de
conflicto de intereses
Los autores declararon no tener
conflictos de intereses de ningún tipo.
Abiri, A., Lavigne, M.,
Rezaei, M., Nikzad, S., Zare, P., Mergny, J. L., & Rahimi, H. R. (2021).
Unlocking G-Quadruplexes as Antiviral Targets. Pharmacological Reviews,
73(3), 897-923. https://doi.org/10.1124/PHARMREV.120.000230
Amrane, S., Jaubert, C.,
Bedrat, A., Rundstadler, T., Recordon-Pinson, P., Aknin, C., Guédin, A.,
De-Rache, A., Bartolucci, L., Diene, I., Lemoi-ne, F., Gascuel, O., Pratviel,
G., Mergny, J. L., & Andreola, M. L. (2022). Deciphering RNA G-quadruplex function
during the early steps of HIV-1 infection. Nucleic Acids Research,
50(21), 12328-12343. https://doi.org/10.1093/NAR/GKAC1030
Artusi, S., Ruggiero, E.,
Nadai, M., Tosoni, B., Perrone, R., Ferino, A., Zanin, I., Xodo, L., Flamand,
L., & Richter, S. N. (2021). Antiviral activity of the g-quadruplex ligand
tmpyp4 against herpes simplex virus-1. Viruses, 13(2). https://doi.org/10.3390/V13020196
Bian, W. X., Xie, Y., Wang,
X. N., Xu, G. H., Fu, B. S., Li, S., Long, G., Zhou, X., & Zhang, X. L.
(2019). Binding of cellular nucleolin with the viral core RNA G-quadruplex
structure suppresses HCV replication. Nucleic Acids Research, 47(1),
56-68. https://doi.org/10.1093/NAR/GKY1177
Biswas, B., Kandpal, M.,
& Vivekanandan, P. (2017). A G-quadruplex motif in an envelope gene
promoter regulates transcription and virion secretion in HBV genotype B. Nucleic
Acids Research, 45(19), 11268-11280. https://doi.org/10.1093/NAR/GKX823
Biver, T. (2022).
Discriminating between Parallel, Anti-Parallel
and Hybrid G-Quadruplexes: Mechanistic Details on Their Binding to Small
Molecules. Molecules, 27(13). https://doi.org/10.3390/MOLECULES27134165
Bohálová, N., Cantara, A., Bartas, M., Kaura,
P., Šťastný, J., Pečinka, P., Fojta, M., Mergny, J. L., & Brázda, V.
(2021). Analyses
of viral genomes for G-quadruplex forming sequences reveal their correlation
with the type of infection. Biochimie, 186, 13-27. https://doi.org/10.1016/J.BIOCHI.2021.03.017
Brázda, V., Valková, N., Dobrovolná, M.,
& Mergny, J. L. (2023). Abundance
of G-Quadruplex Foring Sequences in the Hepatitis Delta Virus Genomes. ACS
Omega, 9(3), 4096-4101. https://doi.org/10.1021/acsomega.3c09288
Butovskaya, E., Heddi, B.,
Bakalar, B., Richter, S. N., & Phan, A. T. (2018). Major G-Quadruplex Form
of HIV-1 LTR Reveals a (3 + 1) Folding Topology Containing a Stem-Loop. Journal
of the American Chemical Society, 140(42), 13654-13662. https://doi.org/10.1021/jacs.8b05332
Caterino, M. &
Paeschke, K. (2022). Action and function of helicases on RNA G-quadruplexes. Methods (San Diego, Calif.), 204, 110-125.
https://doi.org/10.1016/J.YMETH.2021.09.003
Cui, H. & Zhang, L.
(2020). G-Quadruplexes Are Present in Human Coronaviruses Including SARS-CoV-2.
Frontiers in Microbiology, 11. https://doi.org/10.3389/FMICB.2020.567317
Das, P., Winnerdy, F. R.,
Maity, A., Mechulam, Y., & Phan, A. T. (2021). A novel minimal motif for
left-handed G-quadruplex formation. Chemical Communications, 57(20),
2527-2530. https://doi.org/10.1039/D0CC08146A
Datta, A., Pollock, K. J.,
Kormuth, K. A., & Brosh, R. M. (2021). G-Quadruplex Assembly by Ribosomal DNA: Emerging Roles in Disease Pathogenesis
and Cancer Biology. Cytogenetic and Genome Research,
161(6-7), 285-296. https://doi.org/10.1159/000516394
De-Nicola, B., Lech, C. J.,
Heddi, B., Regmi, S., Frasson, I., Perrone, R., Richter, S. N., & Phan, A.
T. (2016). Structure and possible function of a G-quadruplex in the long
terminal repeat of the proviral HIV-1 genome. Nucleic Acids Research,
44(13), 6442-6451. https://doi.org/10.1093/NAR/GKW432
Debbarma, S. & Acharya, P. C. (2022). Targeting G-Quadruplex DNA for Cancer Chemo-therapy. Current Drug Discovery
Technologies,19(3). https://doi.org/10.2174/1570163819666220214115408
Di-Antonio,
M., Ponjavic, A., Radzevičius, A., Ranasinghe, R. T., Catalano, M., Zhang, X., Shen,
J., Needham, L. M., Lee, S. F., Klenerman, D., & Balasubramanian, S.
(2020). Single-molecule visualization of DNA G-quadruplex formation in live
cells. Nature Chemistry, 12(9), 832-837.
https://doi.org/10.1038/S41557-020-0506-4
Dinh, V. T., Loaëc, N.,
Quillévéré, A., Le-Sénéchal, R., Keruzoré, M., Martins, R. P., Granzhan, A.,
& Blondel, M. (2023). The hide-and-seek game of the oncogenic Epstein-Barr
virus-encoded EBNA1 protein with the immune system: An RNA G-quadruplex tale. Biochimie,
214(Pt A), 57-68. https://doi.org/10.1016/j.biochi.2023.07.010
Edwards, A. M., Baric, R.
S., Saphire, E. O., & Ulmer, J. B. (2022). Stopping pandemics before they
start: Lessons learned from SARS-CoV-2. Science, 375(6585), 1133-1139.
https://doi.org/10.1126/SCIENCE.ABN1900
Frasson, I., Nadai, M.,
& Richter, S. N. (2019). Conserved G-Quadruplexes Regulate the Immediate
Early Promoters of Human Alphaherpesviruses. Molecules
(Basel, Switzerland), 24(13).
https://doi.org/10.3390/MOLECULES24132375
Fay, M. M., Lyons, S. M.,
& Ivanov, P. (2017). RNAG-Quadruplexes in
Biology: Principles and Molecular Mechanisms. Journal of molecular biology,
429(14), 2127–2147. https://doi.org/10.1016/j.jmb.2017.05.017
Gellert, M., Lipsett, M.
N., & Davies, D. R. (1962). Helix
formation by guanylic acid. Proceedings of the National Academy of Sciences
of the United States of America (PNAS), 48(12),
2013-2018.https://doi.org/10.1073/pnas.48.12.2013
Gemmill, D. L., Nelson, C.
R., Badmalia, M. D., Pereira, H. S., Kerr, L., Wolfinger, M. T., & Patel,
T. R. (2024). The 3' terminal region of Zika virus RNA contains a conserved
G-quadruplex and is unfolded by human DDX17. Biochemistry and Cell Biology,
102(1), 96-105. https://doi.org/10.1139/BCB-2023-0036
Hargrave, A., Mustafa, A.
S., Hanif, A., Tunio, J. H., & Hanif, S. N. M. (2021). Current Status of
HIV-1 Vaccines. Vaccines, 9(9). https://doi.org/10.3390/VACCINES9091026
Henderson, E., Hardin, C.
C., Walk, S. K., Tinoco, I., &Blackburn, E. H. (1987). Telomeric DNA oligonucleotides form novel intramolecular structures
containing guanine·guanine base pairs. Cell, 51(6), 899-908. https://doi.org/10.1016/0092-8674(87)90577-0
Ji, D., Juhas, M., Tsang,
C. M., Kwok, C. K., Li, Y., & Zhang, Y.
(2021). Discovery of G-quadruplex-forming sequences in SARS-CoV-2. Briefings
in Bioinformatics, 22(2), 1150-1160. https://doi.org/10.1093/BIB/BBAA114
Koonin, E. V., Dolja, V.
V., & Krupovic, M. (2022). The logic of virus evolution. Cell Host and
Microbe, 30(7), 917-929. https://doi.org/10.1016/j.chom.2022.06.008
Kusov, Y., Tan, J.,
Alvarez, E., Enjuanes, L., & Hilgenfeld, R. (2015). A G-quadruplex-binding
macrodomain within the “SARS-unique domain” is essential for the activity of
the SARS-coronavirus replication-transcription complex. Virology, 484,
313-322. https://doi.org/10.1016/J.VIROL.2015.06.016
Lavezzo, E., Berselli, M.,
Frasson, I., Perrone, R., Palù, G., Brazzale, A. R., Richter, S. N., &
Toppo, S. (2018). G-quadruplex forming sequences in the genome of all known
human viruses: A comprehensive guide. PLoS Computational Biology,
14(12), e1006675. https://doi.org/10.1371/journal.pcbi.1006675
Lyu, K., Chow, E. Y. C.,
Mou, X., Chan, T. F., & Kwok, C.K. (2021). RNA G-quadruplexes (rG4s):
genomics and biological functions. Nucleic Acids Research, 49(10),
5426-5450. https://doi.org/10.1093/NAR/GKAB187
Majee, P., Kumar-Mishra, S., Pandya, N., Shankar,
U., Pasadi, S., Muniyappa, K., Nayak, D., & Kumar, A. (2020). Identification and
characterization of two conserved G-quadruplex for-ming motifs in the Nipah
virus genome and their interaction with G-quadruplex specific ligands. Scientific
Reports 10(1), 1-12. https://doi.org/10.1038/s41598-020-58406-8
Marušič, M., Hošnjak, L.,
Krafčikova, P., Poljak, M.,Viglasky, V., & Plavec,
J. (2017). The effect of single nucleotide polymorphisms in G-rich regions of
high-risk human papillomaviruses on structural diversity of DNA. Biochimica
et Biophysica Acta. General Subjects, 1861(5 Pt B), 1229-1236. https://doi.org/10.1016/J.BBAGEN.2016.11.007
Marušič, M. & Plavec,
J. (2019). Towards understanding of polymorphism of the g-rich region of human
papillomavirus type 52. Molecules, 24(7). https://doi.org/10.3390/MOLECULES24071294
Meier-Stephenson, V.,
Badmalia, M. D., Mrozowich, T., Lau, K. C. K., Schultz, S. K., Gemmill, D. L.,
Osiowy, C., van Marle, G., Coffin, C. S., & Patel, T. R. (2021).
Identification and characterization of a G-quadruplex structure in the pre-core
promoter region of hepatitis B virus covalently closed circular DNA. The
Journal of Biological Chemistry, 296.
https://doi.org/10.1016/J.JBC.2021.100589
Pont, I. &
González-García, J. (2022). Beyond the double
helix: Structure of DNA G-quadruplexes. Metode Science Studies Journal,
(12), 6-13. https://doi.org/10.7203/metode.12.16505
Piekna-Przybylska, D.,
Sullivan, M. A., Sharma, G., & Bambara, R. A. (2014). U3 region in the
HIV-1 genome adopts a G-quadruplex structure in its RNA and DNA sequence. Biochemistry,
53(16), 2581-2593. https://doi.org/10.1021/BI4016692
Puig-Lombardi, E. &
Londoño-Vallejo, A. (2020). A guide to computational methods for G-quadruplex
prediction. Nucleic Acids Research, 48(1), 1-15. https://doi.org/10.1093/NAR/GKZ1097
Ravichandran, S., Kim, Y.
E., Bansal, V., Ghosh, A., Hur, J., Subramani, V. K., Pradhan, S., Lee, M. K.,
Kim, K. K., & Ahn, J. H. (2018). Genome-wide analysis of regulatory
G-quadruplexes affecting gene expression in human cytomegalovirus. PLoS
Pathogens, 14(9). https://doi.org/10.1371/JOURNAL.PPAT.1007334
Ruggiero, E. & Richter,
S. N. (2020). Viral G-quadruplexes: New frontiers in virus pathogenesis and
antiviral therapy. Annual Reports in Medicinal Chemistry, 54, 101-131. https://doi.org/10.1016/BS.ARMC.2020.04.001
Ruggiero, E., Zanin, I.,
Terreri, M., & Richter, S. N. (2021). G-quadruplex targeting in the fight
against viruses: An update. International Journal of Molecular Sciences,
22(20). https://doi.org/10.3390/IJMS222010984
Satkunanathan, S., Thorpe,
R., & Zhao, Y. (2017). The function of DNA binding protein nucleophosmin in
AAV replication. Virology, 510, 46-54. https://doi.org/10.1016/J.VIROL.2017.07.007
Sato, K. & Knipscheer,
P. (2023). G-quadruplex resolution: From
molecular mechanisms to physiological relevance. DNA Repair, 130. https://doi.org/10.1016/j.dnarep.2023.103552
Summers, P. A., Lewis, B. W.,
Gonzalez-Garcia, J., Porreca, R. M., Lim, A. H. M., Cadinu, P., Martin-Pintado,
N., Mann, D. J., Edel, J. B., Vannier, J. B., Kuimova, M. K., & Vilar, R.
(2021). Visualising
G-quadruplex DNA dynamics in live cells by fluorescence lifetime imaging
microscopy. Nature Communications, 12(1), 1-11. https://doi.org/10.1038/s41467-020-20414-7
Teng, F. Y., Jiang, Z. Z.,
Guo, M., Tan, X. Z., Chen, F., Xi, X. G., & Xu, Y. (2021). G-quadruplex
DNA: a novel target for drug design. Cellular and Molecular Life Sciences,
78(19-20), 6557-6583. https://doi.org/10.1007/S00018-021-03921-8,
Tlučková, K., Marušič, M., Tóthová, P., Bauer, L.,
Šket, P., Plavec, J., & Viglasky, V. (2013). Human papillomavirus G-quadruplexes. Biochemistry,
52(41), 7207-7216. https://doi.org/10.1021/BI400897G
Tuesuwan, B., Kern, J. T.,
Thomas, P. W., Rodriguez, M., Li, J., David, W. M., & Kerwin, S. M. (2008).
Simian virus 40 large T-antigen G-quadruplex DNA helicase inhibition by
G-quadruplex DNA-interactive agents. Biochemistry, 47(7), 1896-1909. https://doi.org/10.1021/BI701747D
Wang, S. R., Min, Y. Q.,
Wang, J. Q., Liu, C. X., Fu, B. S., Wu, F., Wu, L. Y., Qiao, Z. X., Song, Y.
Y., Xu, G. H., Wu, Z. G., Huang, G., Peng, N. F., Huang, R., Mao, W. X., Peng,
S., Chen, Y. Q., Zhu, Y., Tian, T., …, & Zhou, X. (2016a). A highly
conserved G-rich consensus sequence in hepatitis C virus core gene represents a
new anti–hepatitis C target. Science
Advances, 2(4), e1501535. https://doi.org/10.1126/SCIADV.1501535
Wang, S. R., Zhang, Q. Y.,
Wang, J. Q., Ge, X. Y., Song, Y. Y., Wang, Y. F., Li, X. D., Fu, B. S., Xu, G.
H., Shu, B., Gong, P., Zhang, B., Tian, T., & Zhou, X. (2016b). Chemical
Targeting of a G-Quadruplex RNA in the Ebola Virus L Gene. Cell Chemical
Biology, 23(9), 1113-1122. https://doi.org/10.1016/J.CHEMBIOL.2016.07.019
Yan, M. P., Wee, C. E.,
Yen, K. P., Stevens, A., & Wai, L. K. (2023). G-quadruplex ligands as
therapeutic agents against cancer, neurological disorders and viral infections.
Future Medicinal Chemistry, 15(21), 1987-2009. https://doi.org/10.4155/fmc-2023-0202
Zareie, A. R., Dabral, P.,
& Verma, S. C. (2024). G-Quadruplexes in the Regulation of Viral Gene
Expressions and Their Impacts on Controlling
Infection. Pathogens, 13(1). https://doi.org/10.3390/PATHOGENS13010060